Les Annélides
Les Annélides sont constitués d'une série de segments provenant d'un développement embryonnaire bien précis (voir la partie « Développement de la larve trochophore). Nous y avons vu apparaître la métamérisation caractéristique des Annélides et comment elle détermine la structure globale de l’animal.
Nous verrons concrètement comment cette structure métamérisée fonctionne chez un Annélide adulte. Nous illustrerons ceci lors des séances de travaux pratiques avec l'étude d'un animal d'un intérêt écologique primordial : le Ver de Terre.
Enfin, la diversité de l'Embranchement sera évoquée. Nous y observerons des adaptations progressives à des modes de vies différents à travers l’étude de quelques exemples.
Fiche récapitulative
- Métazoaires, triploblastiques, coelomates ;
- Symétrie bilatérale ;
- Protostomiens ;
- Métamérisés ;
- Corps en 3 parties: tête, soma, telson ;
- Tube digestif complet formé de régions spécialisées ;
- Système circulatoire fermé ;
- Système d’excrétion différencié (métanéphridies) ;
- Système nerveux formé d’un ganglion antérieur supraoesophagien et d’une chaîne nerveuse ganglionnaire ventrale ;
- Système reproducteur pouvant se combiner au bourgeonnement ;
- Marins, dulçaquicoles, ou terrestres ;
- Libres ou ectoparasites.

1. Présentation du groupe
Les Annélides sont des vers annelés, comme l'indique leur nom. Leur taille peut varier entre 0,5 mm et 3 m de longueur. La plupart des Annélides vivent en eau de mer. Quelques-uns ont conquis la terre, comme le Ver de terre, que vous voyez sur cette photo, et les eaux douces. Vous aurez l’occasion d’en observer différentes espèces lors des travaux pratiques.

Nereis
Cet autre Annélide appartient au genre Nereis, dont la plupart des espèces vivent en eau de mer. Nereis fouit les sédiments marins et se déplace aussi à leur surface.

Sangsue
Et voici une Sangsue en train de déjeuner. Elle représente un troisième grand groupe d'Annélides.

2. Exemple-type : le ver de terre (Lumbricus terrestris)
Anecdote : le saviez-vous ?
Les vers de terre furent qualifiés de « premiers laboureurs » par Darwin, qui leur consacra tout un livre. En effet, ils ont une fonction essentielle d'aération et de structuration des sols. Ils contribuent fortement à leur fertilité, en transformant les déchets organiques (feuilles mortes, défécations d'animaux, etc.) en composés directement assimilables par les plantes. Les réactions que suscitent les vers de terre sont diverses: ils ont une bonne réputation auprès des pêcheurs et des jardiniers, provoquent la répugnance du public, l'intérêt des agronomes, ...
Lorsque le sol est humide, on peut vérifier la présence des vers de terre dans chaque bêchée de terre, que ce soit en forêt, dans une prairie, un jardin ou un champ. Cette omniprésence en région tempérée traduit une réussite écologique étonnante.

Anecdote
Comme vous le voyez sur cette publicité, les vers de terre peuvent être utilisés comme agents de compostage par tout un chacun. A ce titre, et aussi par d'autres aspects bénéfiques dont nous reparlerons, les vers de terre occupent une position irremplaçable dans l’économie actuelle et de demain. Encore faut-il que l'Homme en soit conscient et qu'il connaisse mieux leur fonction exacte.

2.1. Examen externe
L’animal est long, cylindrique, annelé : chaque anneau correspond à un métamère. Il présente une symétrie bilatérale et un axe antéro-postérieur. La face dorsale est plus pigmentée que la face ventrale.

Le ver de terre
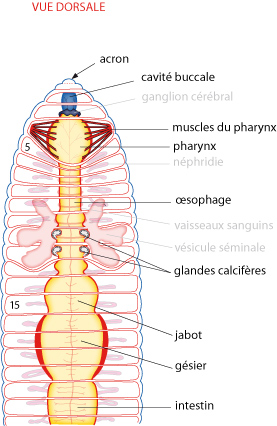
Sur cette figure on observe le Lombric par sa face dorsale et son extrémité antérieure, l’acron se trouvant tout à gauche et on peut facilement compter les segments.
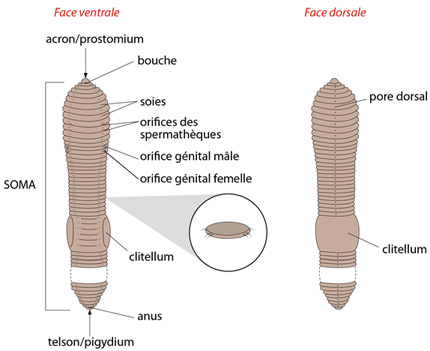
Le soma
Nous le regardons maintenant plus en détail par les faces ventrale et dorsale. Le soma est constitué de plus d'une centaine de segments (ou métamères), précédés d'un premier segment nommé acron (ou prostomium) et suivis d'un telson (ou pygidium) à l'arrière. Tous les métamères semblent identiques... En fait, ce n'est pas exact : déjà à l'examen externe, on aperçoit que tous les segments n’ont pas la même largeur. Ainsi, les premiers segments antérieurs sont plus larges que ceux qui les suivent. Ceci correspond à des différenciations spéciales du tube digestif dans cette région; nous le verrons bientôt. De plus, au moment de la reproduction, le tégument présente un renflement du 33ème au 38ème métamères, le clitellum, qui a l'air d'être posé sur le dos du ver comme une selle.
Les 4 segments antérieurs du ver
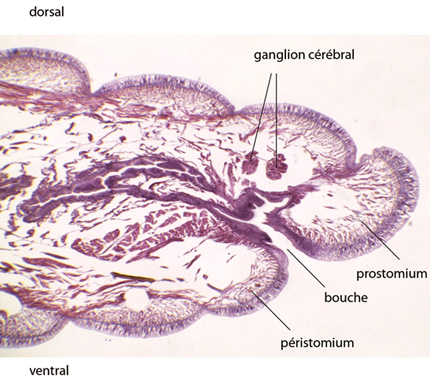
Juste derrière l'acron s'ouvre la bouche, devant un segment que l’on appelle péristomium, tandis que l'anus se situe entre le dernier métamère du soma et le telson.
Paires de soies
Chaque métamère porte latéro-ventralement 4 paires de soies, disposées comme vous le voyez sur la figure. On perçoit facilement les soies en passant son doigt sur le corps, d’arrière en avant, comme vous le constaterez en travaux pratiques.
Le Lombric dispose d'une série complexe d'orifices pairs liés au système reproducteur. Attention leur emplacement varie d'une espèce de Lombric à l'autre.
Sur la face ventrale de Lumbricus terrestris :
- les ouvertures des spermathèques (réceptacles séminaux) sont en arrière des 9ème et 10ème segments ;
- les orifices génitaux femelles sont sur le 14ème segment ;
- les orifices génitaux mâles sont sur le 15ème segment.
Dorsalement, un pore dorsal par métamère permet l'évacuation rapide du liquide cœlomique, en cas de besoin. Nous n'avons pas représenté les deux orifices excréteurs (les néphridiopores) situés sur chaque métamère, en position ventro-latérale: ils sont très petits. De plus, ces orifices sont fermés par des muscles sphincters qui lorsqu’ils sont contractés rendent l’observation des différents pores très difficile.
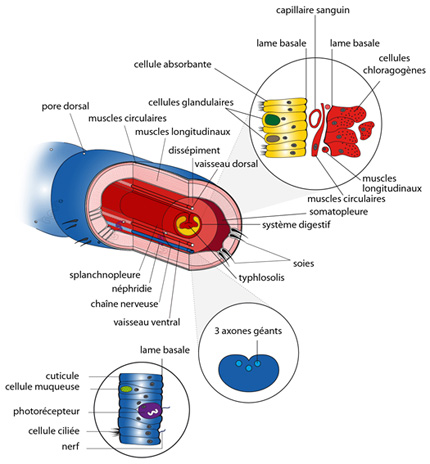
2.2. Examen interne
L’examen interne pourra se faire durant le TP sur la dissection du Lombric et grâce aux coupes microscopiques mises à votre disposition. Quelle que soit la région du corps observée, on remarque qu’il y a concordance entre l’annellation externe et la métamérisation interne. Cette image, reprise aussi dans votre syllabus, représente une vue éclatée du Lombric, que nous allons compléter petit à petit.
2.2.1. Tégument et épiderme
Nous allons commencer par la paroi externe du Lombric : l'épiderme qui est un dérivé ectodermique. Il n'est interrompu que par le passage des soies et les pores. Il sécrète une cuticule protectrice. Voyez le schéma détaillé des cellules épidermiques en cliquant sur le tégument du Lombric de la figure. L'épiderme contient de nombreuses cellules muqueuses et des cellules sensorielles de divers types: des cellules ciliées tactiles, des chémorécepteurs et des photorécepteurs. Le renflement du clitellum est dû à une multiplication locale des cellules muqueuses.
Le mucus sécrété par le tégument, composé ici de l’épiderme, aide le ver de terre à glisser dans les galeries qu'il creuse ; le mucus renforce aussi les parois de ces galeries en séchant quand le ver est passé. Le Lombric ne possède pas d'yeux à proprement parler: dans l’obscurité de l’humus et des galeries, ils lui seraient inutiles. Mais son tégument est pourvu de cellules photosensibles réparties sur tout le corps, un peu plus densément sur la tête. Ainsi, le Lombric peut détecter la lumière et la fuir, car les rayons ultraviolets peuvent le tuer, malgré les pigments de protection qui lui donnent sa coloration brune. Par contre d’autres espèces d’Annélides possèdent des yeux simples sur la tête, surtout parmi les Polychètes que l’on verra par la suite.
Exercice
Définir phototactisme négatif :
Définir chimiotactisme :
2.2.2. Système locomoteur
Musculature périphérique : les différents éléments d’origine mésodermique, dont les muscles, apparaissent en rouge.
Située juste sous le tégument, la musculature périphérique, dérivée de la somatopleure mésodermique, comporte une assise circulaire externe et une assise longitudinale interne, dont la monotonie est rompue par les petits muscles obliques servant à faire se mouvoir les soies. La musculature longitudinale est divisée en quatre champs, un dorsal, un ventral et deux latéraux. Un mince feuillet somatopleural tapisse la surface externe des muscles.
Le système digestif est entouré d’une splanchnopleure. Au cours du développement, la splanchnopleure donne la musculature péri-intestinale ainsi que les cellules chloragogènes et potentiellement le système circulatoire.
Les coelomes
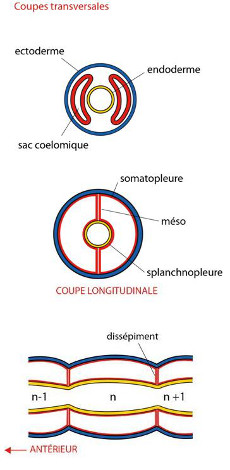
Rappelons que chaque segment ou métamère contient une paire de cœlomes symétriques situés de part et d’autre du tube digestif, et qui forment une double paroi, le méso ou mésentère. On retrouve un mésentère dorsal et un mésentère ventral. Les cœlomes entrent également en contact entre métamères successifs et forment des dissépiments.
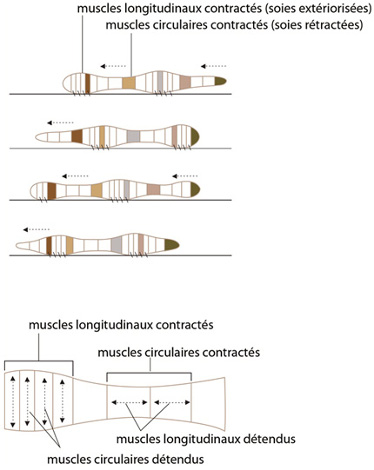
Locomotion
Lorsque le ver se déplace, le liquide cœlomique fonctionne en tant que squelette hydrostatique, sur lequel agissent les muscles pour modifier la forme des métamères et permettre le déplacement. Le ver se déplace de la façon suivante: dans la partie antérieure du corps, la contraction des muscles circulaires comprime le liquide cœlomique dans les métamères concernés, qui s’allongent. Les soies sont rétractées. En même temps, la contraction des muscles longitudinaux dans les métamères plus postérieurs y comprime le liquide cœlomique et provoque leur élargissement. Leurs soies s’ancrent dans le substrat. Ils se sont rapprochés des métamères antérieurs. Puis, ces métamères antérieurs contractent leur musculature longitudinale ; leurs soies s’ancrent. Ces vagues de contraction-relâchement des musculatures circulaires et longitudinales se propagent le long du ver et le font avancer.
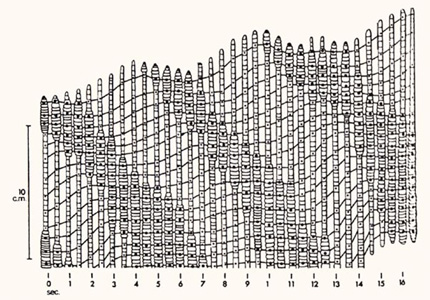
Déplacement du ver de terre
Donc, l’élargissement ou l’allongement se succèdent entre métamères. Grâce à ce squelette hydrostatique, une très forte puissance est disponible; l'animal peut creuser le sol par son extrémité antérieure (d'ailleurs, l'acron est fort résistant); il peut aussi se mouvoir très rapidement dans une galerie qui est déjà creusée.
2.2.3. Système digestif - Structure générale
Sur la vue éclatée du Lombric nous complétons maintenant les éléments du système digestif (cliquez sur système digestif de la figure). Chez les Annélides, la plupart des structures dérivées de l’ectoderme et du mésoderme sont atteintes par la métamérisation: muscles circulaires et longitudinaux, système excréteur formés de paires de métanéphridies, système circulatoire comportant des vaisseaux circulaires, ganglions du système nerveux. Par contre, les structures dérivées de l’endoderme (en jaune sur la figure) ne sont pas répétées: le système digestif est non métamérisé.
Dans un métamère typique, la forme du tube digestif n’est compliquée que par une invagination dorsale de la paroi intestinale dans la lumière. C’est le typhlosolis, qui permet d’augmenter la surface absorbante du tube digestif.
Cliquez sur l’intérieur du tube digestif de la figure afin d’observer les cellules qui constituent la paroi. L’épithélium dérivé de l’endoderme, en jaune, comprend des cellules absorbantes bien sûr, munies de cils, et des cellules glandulaires, déversant des enzymes digestifs et du mucus dans la lumière. Les nutriments absorbés diffusent en petite partie vers la cavité cœlomique, mais l’essentiel est prélevé dans le tissu conjonctif situé sous la lame basale de l'épithélium, riche en vaisseaux capillaires qui véhiculent les nutriments dans le système circulatoire. Extérieurement au réseau capillaire, on trouve des muscles circulaires, puis des muscles longitudinaux. Enfin, de grandes cellules appelées chloragogènes formant la splanchnopleure. Ces cellules sont le siège d'une partie importante du métabolisme des glucides et des protides ; elles remplissent donc certaines fonctions du foie des Vertébrés. Elles stockent les graisses. En plus, elles interviennent dans la formation d'urée, d’acide urique et d'autres produits voués à l'excrétion. Une fois chargées, elles se détachent et tombent dans la cavité cœlomique. La musculature intestinale et les capillaires proviennent aussi de la différenciation de la splanchnopleure embryonnaire, et sont donc d’origine mésodermique.
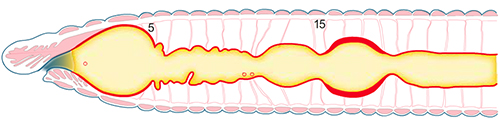
Tube digestif antérieur
Ce que nous venons de voir sur la figure, est la structure du tube digestif d’un métamère plutôt postérieur. La partie antérieure du tube digestif présente une belle spécialisation qui sera visible lors de la dissection ou à travers l’observation de coupes microscopiques longitudinales.
La bouche est disposée ventralement au niveau du 2ième segment qui a fusionné avec l'acron. La cavité buccale est tapissée par un épithélium d’origine ectodermique. En arrière de la bouche, se trouve un pharynx tapissé d'endoderme comme la suite du tube digestif, et à la paroi musculaire très épaissie. Puis un oesophage conduit à un jabot, où les matières ingérées peuvent s'accumuler avant de passer dans un gésier musclé, où elles sont broyées.
Le pharynx
Des faisceaux musculaires du pharynx rejoignent la paroi du corps. Le pharynx est donc capable d'aspiration ; il force l'ingestion de la terre, des débris végétaux et autres sources de nourriture.
Différentes glandes sont annexées à la portion antérieure du tube digestif. Des glandes salivaires déversent leurs sécrétions enzymatiques dans le pharynx ; d’autres glandes appelées calcifères sont connectées à l'oesophage: elles interviennent dans la régulation ionique plutôt que dans la digestion. En effet, le Lombric est exposé à une alcalinisation excessive de son sang étant donné un régime alimentaire riche en calcium. Les glandes calcifères combinent le CO2 du sang avec le calcium, sous forme de carbonate insoluble. Le carbonate de calcium sera évacué dans la lumière de l'œsophage et il transitera tout au long du tube digestif sans être réabsorbé. C'est une solution originale que propose le Lombric.
Exercice
Comment se fait-il que la cavité buccale soit tapissée d'ectoderme ?
Au vu de la structure cellulaire caractéristique de l’ectoderme, y voyez-vous un avantage ?
2.2.4. Système respiratoire
L'apport alimentaire étant assuré, comment le Lombric se procure-t-il l'oxygène nécessaire à son métabolisme et élimine-t-il le CO2 (celui qui n'est pas combiné dans les glandes calcifères) ? Simplement par diffusion des gaz à travers son tégument. Pour cela, il dispose de réseaux capillaires disposés sous l'ectoderme. Par ailleurs, le sang du Lombric contient de l'hémoglobine, qui donne au sang sa couleur rouge.
Exercice
Chez les Mammifères, l’hémoglobine est contenue dans des cellules : les globules rouges.
Chez le Lombric, elle est en solution dans le sang. Quels sont les avantages et les inconvénients de ces deux modalités ?
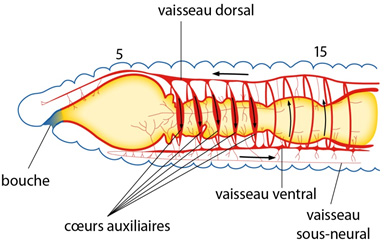
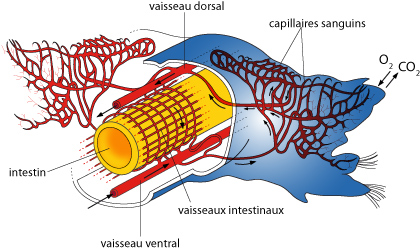
2.2.5. Système circulatoire
Le système circulatoire suit de très près le plan de base que nous avons évoqué précédemment. Le sang reste entièrement enfermé dans des vaisseaux; le système est « fermé ». Ventralement, on retrouve d’ailleurs un vaisseau ventral et un vaisseau sous-neural (figure AN.2.13). Les vaisseaux dorsal et ventral sont connectés l'un à l'autre par des vaisseaux circulaires, au niveau de chaque dissépiment (figure AN.2.13). Le vaisseau ventral est suspendu au tube digestif par le mésentère ventral. Le vaisseau ventral se ramifie en branches puis en capillaires dans le tégument, l'intestin et les autres organes, puis le sang est ramené aux plus gros vaisseaux principaux. Le sang est principalement propulsé par les contractions du vaisseau dorsal. Chez le Lombric, cette propulsion est aidée par des cœurs auxiliaires : il y en a 5 paires disposées autour de l'œsophage (figure AN.2.13). Le sang circule vers l'avant dans le vaisseau dorsal, et vers l'arrière dans les vaisseaux ventraux. Notez à la face ventrale le vaisseau sous-neural, qui assure l’irrigation de la chaîne nerveuse.
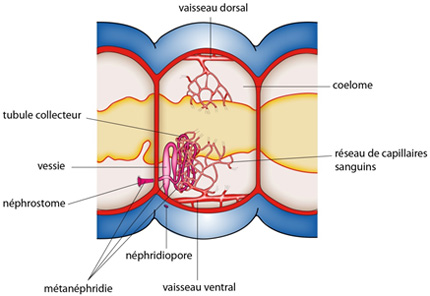
2.2.6. Système excréteur
Comme la grande majorité des Annélides, le Lombric possède typiquement une paire de néphridies (aussi appelées métanéphridies) par métamère. Le système excréteur a une double fonction: éliminer les déchets du métabolisme et maintenir l'équilibre osmotique. La néphridie du Lombric propose déjà des spécialisations intéressantes, mais elle n'est pas encore parfaitement adaptée à la vie terrestre. Chaque néphridie comporte un néphrostome cilié qui s'ouvre dans le métamère précédent et nettoie le coelome du métamère précédent.
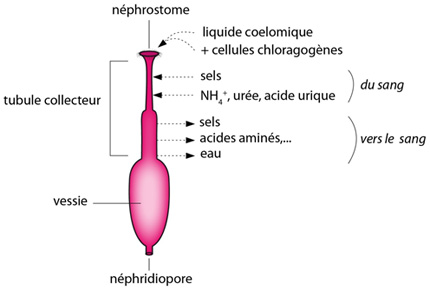
La néphridie du lombric
Par le néphrostome cilié, le liquide cœlomique, charriant de vieilles cellules chloragogènes bourrées de déchets métaboliques, s'engouffre dans le tube néphridien au niveau du tubule collecteur. Là, dans une première région, les capillaires sanguins irriguant la paroi vont rejeter dans la lumière du tube quelques sels, et surtout des produits terminaux du catabolisme azoté (de l'urée, de l'acide urique). Plus loin, des produits intéressants comme des acides aminés, des sucres, des sels et de l’eau seront récupérés et repris dans des capillaires sanguins. Avant d'être évacuée à l'extérieur par le néphridiopore, l'urine peut être stockée quelques temps dans la vessie terminale de la néphridie.
Puisqu'il court le risque de perdre de l'eau par son tégument et par ses néphridies, le Lombric est obligé de rester dans des terrains relativement humides. Ceci ne veut pas dire que les sols plus secs sont invivables pour les Annélides. Certaines espèces ont inventé une solution originale : elles ont dévié leurs néphridies, qui ne s'ouvrent plus directement à l'extérieur par un néphridiopore, mais débouchent dans l'intestin ! Ainsi, ils profitent des capacités de réabsorption de l'épithélium intestinal, très performant à ce point de vue.
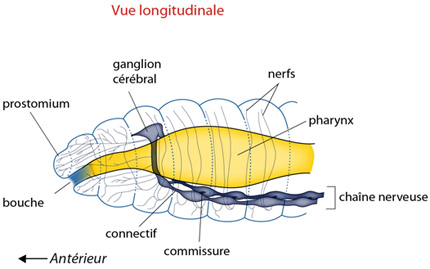
2.2.7. Système nerveux
L'activité du Lombric est coordonnée par un système nerveux campé sur le schéma général vu précédemment: un cerveau antérieur, au-dessus de la cavité buccale, et une double chaîne ganglionnaire ventrale, fusionnée sur la ligne médiane. Le système nerveux est métamérisé et on retrouve une paire de ganglions par métamère.
Les axones
Comme vous pourrez le constater facilement sur la coupe transversale au microscope, et sur la figure en cliquant sur la chaine nerveuse, le Lombric dispose d'axones géants. Un groupe de trois axones est bien visible dans la région dorsale de la chaîne ganglionnaire. Ils s'étendent d'un bout à l'autre du ver et permettent la transmission très rapide d'influx nerveux. Ainsi, lorsque le Lombric émerge dans l'humidité du soir, il ne s'éloigne jamais bien loin de l'orifice de sa galerie; il y laisse même enfoncée son extrémité postérieure. Si un danger survient, un oiseau prédateur par exemple, les axones géants transmettent immédiatement l'alerte à tous les neurones moteurs de la musculature et le Lombric se réfugie instantanément dans son domicile. Au sein de la chaîne nerveuse, les corps cellulaires des neurones occupent une position périphérique et les axones forment la partie centrale.
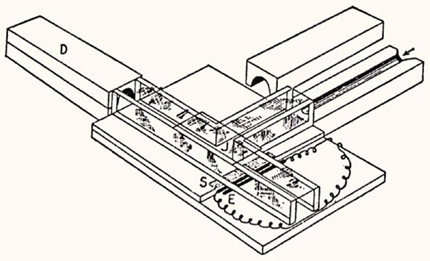
Lumbricus : anecdote
Comme les vers de terre sont économiquement très intéressants et qu’ils sont abondants, on leur a fait subir de multiples expériences, les plus diverses. On a même essayé de savoir non pas s'ils étaient intelligents, mais s'ils pouvaient apprendre. Voici le labyrinthe de YERKES. Le ver est placé à l'entrée de ce labyrinthe en forme de T. A gauche, danger il y a une plaque de verre qui griffe son tégument et des électrodes qui lui donnent des décharges ; à droite, c'est le paradis : un beau trou noir plein de feuilles mortes appétissantes. Aux premiers essais, le ver emprunte aléatoirement les deux voies. Après quelques semaines d'essais, il va directement à droite dans 90% des cas. Mais si on suspend l'apprentissage, il a tout oublié après quelques jours !
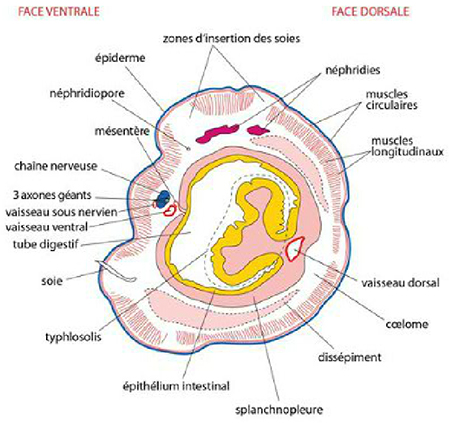
Les parties du lombric
Avant de passer au dernier système, retrouvez toutes les parties du lombric sur cette coupe transversale.
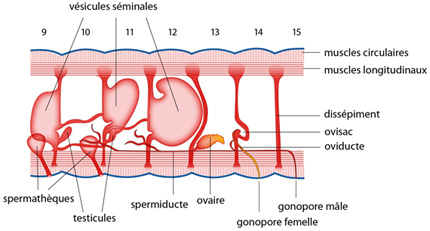
2.2.8. Système reproducteur et régénération
Contrairement à un préjugé tenace, un Lombric coupé en deux par un coup de bêche ne peut jamais donner deux individus viables! En revanche, la capacité de régénération des vers de terre est étonnante. Ainsi, un individu dont l'extrémité postérieure est sectionnée pourra la régénérer. Cependant, la capacité de régénération varie suivant l'axe antéro-postérieur: la queue sera plus facile à régénérer que la tête.
Nous verrons que d'autres espèces d'Annélides profitent en quelque sorte de ces capacités de régénération pour pratiquer une reproduction asexuée.
Chez le Lombric, le mode habituel de reproduction est sexué et est précédé d'un accouplement. Le Lombric est hermaphrodite et il dispose d'un système génital complexe qui varie d’une espèce de Lombric à une autre. A titre informatif, voici la disposition précise du système reproducteur, ainsi que les détails de fonctionnement de l’appareil reproducteur.
Deux paires de spermathèques s'ouvrent à la jonction des métamères 9-10 et 10-11. Ce sont des poches où sont entreposés momentanément les spermatozoïdes du partenaire. Les testicules sont disposés par paires sur les dissépiments antérieurs des métamères 10 et 11, et font saillie dans la cavité cœlomique ; ils restent petits, car ils ne font que produire des spermatogonies, libérées dans le cœlome, et captées par des vésicules séminales qui garnissent le dissépiment qui lui fait face ; la spermatogenèse se déroule dans ces vésicules séminales, et lors de la copulation, les spermatozoïdes mûrs se retrouvent à nouveau dans le cœlome. Ils s'engouffrent dans les pavillons de néphridies modifiées en spermiductes. Les spermatozoïdes transitent le long de ces conduits, vers l'arrière, pour déboucher aux deux pores génitaux mâles, sur le 15ème segment.
La formation des gamètes femelles est similaire. La paire d'ovaires fait hernie sur la cloison antérieure du 13ème métamère, et les ovogonies vont mûrir dans l'ovisac d'en face, creusé dans la cloison postérieure. Les œufs seront happés dans la cavité cœlomique par deux néphridies modifiées qui s'ouvrent à l’extérieur sur le segment n°14.

Reproduction du lombric
Par un beau soir d'été, deux Lombrics s'accouplent. Amoureux, mais prudents, chacun est sorti de sa galerie mais il y laisse sa partie postérieure. Ainsi, si un importun survient, il pourra y rentrer à toute vitesse grâce à ses axones géants. Ils se sont reconnus et disposent tête-bêche leur partie antérieure, ventre à ventre ; ils s'accrochent par leurs soies génitales (Figure AN.2.19).
C'est ici que le clitellum entre en jeu. Les Lombrics s'arrangent pour que cet épaississement muqueux de l'épiderme se situe au niveau des spermathèques du partenaire. Le clitellum sécrète du mucus, qui durcit à l'air. L'union est ainsi renforcée par un double manchon muqueux. Les spermatozoïdes sont émis aux pores génitaux du 15ème métamère. Par un périple complexe à la surface des conjoints, ils finissent par gagner les spermathèques de l'autre.
Avec toute cette explication, on ne sera plus étonné de savoir que la copulation des vers dure plusieurs heures... Et ce n'est pas fini.
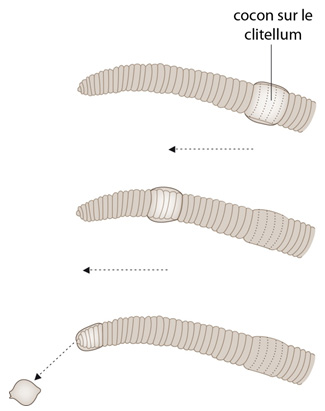
Le cocon
Les conjoints se séparent, mais la fécondation n'a pas encore eu lieu. Le lendemain ou quelques jours plus tard, le clitellum sécrète un cocon annulaire résistant. Le Lombric fait glisser ce cocon vers l'avant ; au passage devant le 14ème métamère, les œufs mûrs sont déposés dans le cocon, le cocon poursuit son chemin. Au passage devant les métamères 10 et 11, les spermathèques sont comprimées et des spermatozoïdes du partenaire, qui attendaient, sont libérés dans le cocon. La fécondation a enfin lieu. Le cocon est expulsé à l'avant du ver, les bouts sont fermés. Les cocons ressemblent à de minuscules citrons, qui contiennent des oeufs. Un seul œuf va s'y développer, aux dépens de ses frères, et au bout de 3 mois environ, un jeune Lombric complet en sortira.
Les spermatozoïdes déposés par le partenaire dans les spermathèques suffiront à ensemencer plusieurs cocons, formés au rythme d'un tous les 4-5 jours.
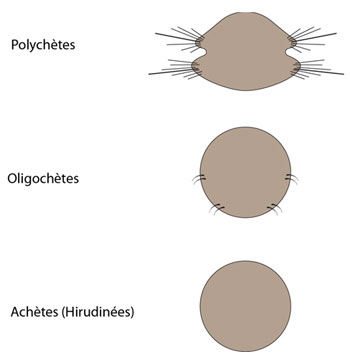
Les 3 clades
Nous avons vu comment est construit et fonctionne un Annélide, précisément le ver de terre. Voyons à présent la diversité de cet Embranchement. L'Embranchement des Annélides se subdivise en 3 clades: les Polychètes, les Oligochètes et les Achètes aussi appelés « Hirudinées ». «-chète» provient de la racine grecque signifiant «soie»; « Poly » veut dire « beaucoup »: les Polychètes ont «beaucoup» de soies. « Oligo » veut dire «peu»: les Oligochètes ont peu de soies et le Ver de terre était l’un d’entre eux. « A » veut dire «sans»: les Achètes n'ont pas de soies du tout. Les schémas de la figure AN.3.1 représentent des coupes transversales simplifiées à l'extrême illustrant ceci.
Précisons ici, avant d’approfondir la diversité de ces groupes, les caractères dérivés propres (« autapomorphies ») des Annélides. Il y a une certaine homogénéité dans le plan d’organisation de tous les Annélides et deux caractères propres sont importants à citer ici: ce sont des animaux fondamentalement métamérisés et ce sont les seuls protostomiens (avec les mollusques céphalopodes) à disposer d’un système circulatoire fermé.
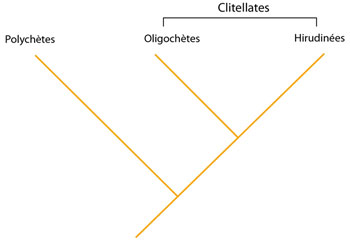
L’évolution des Annélides
L’évolution des Annélides est représentée par le cladogramme de la figure AN.3.2. Les Annélides les plus primitifs sont les Polychètes, puis apparaissent les Oligochètes que l’on regroupe avec les Hirudinées sous le vocable de « Clitellates » : ils possèdent un clitellum. Nous reviendrons sur cette évolution à la fin de ce chapitre.

3.1. Polychètes
La plupart des polychètes sont marins. Les formes libres ou errantes se caractérisent par une tête bien distincte et la présence de parapodes. Les formes sédentaires vivent à l’intérieur de tubes. Voyons quelques exemplaires.
Voici deux représentants de Nereis diversicolor. C'est une espèce très répandue sur les côtes de la Mer du Nord, de la Manche, de l'Atlantique, de la Méditerranée. Elle apprécie la vase, le sable, la boue des estuaires, mais elle est aussi capable de nager. Elle s'accommode aussi bien de l'eau saumâtre que des eaux salées des marais salants. Ce ver mesure une dizaine de centimètres de long et n'est pas plus gros qu'un crayon. Il est de couleurs variées : jaune, rouge, orange, verdâtre.
Il s’agit donc d’un Polychète, un Annélides pourvu de nombreuses soies. Chaque métamère est garni à gauche et à droite de petites excroissances, les parapodes, et (ça, c'est moins visible) dans chaque parapode sont enfoncées de nombreuses soies. On observe également que la partie antérieure comprend une tête bien distincte.

Métamère de polychète
Observons le montage d'un métamère isolé du reste du corps à la figure AN.3.4. Le métamère est légèrement aplati ventralement et présente des parapodes sur les côtés. Les parapodes permettent au ver de nager, ou de creuser des galeries dans le sédiment. Les acicules sont de volumineuses soies internes, visibles en noir sur la photo. A sa base, l’acicule s'insère dans les muscles destinés à mouvoir les parapodes. Vous voyez également d'autres soies, plus petites.
Le ver Nereis se meut en serpentant. Ainsi, en milieu aquatique, les parapodes représentent un soutien et rendent efficaces les mouvements locomoteurs des muscles de la paroi du corps.
Les parapodes
Les parapodes participent aussi à une autre fonction liée à la vie aquatique : la respiration. C'est que la concentration d'oxygène dans l'eau est faible, comparée aux 21 % d’O2 que l'on trouve dans l'air ! Les parapodes offrent un accroissement de la surface tégumentaire, sous laquelle peuvent s'étaler des réseaux de capillaires sanguins responsables des échanges gazeux avec le milieu extérieur (Figure AN.3.5). Chez de nombreux Annélides marins, les parapodes porteront des ébauches de branchies.

La tête de Nereis
Quand le pêcheur ramasse au bord de la mer un Nereis pour l'accrocher à son hameçon, il doit le regarder avec soin et saisir l'animal juste en arrière de la tête, autrement il se fera mordre.
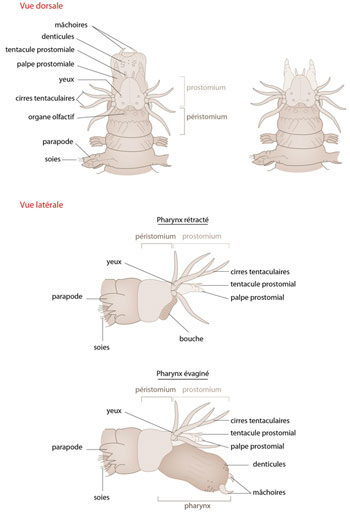
La tête de Nereis
Voici la tête de Nereis, vue dorsalement et latéralement (figure AN.3.8). L'acron/prostomium est fort bien développé et porte de nombreuses structures sensorielles : des tentacules, des palpes, des fossettes ciliées olfactives et 4 yeux ; ce sont les 4 petites taches noires. Le deuxième segment ou péristomium est garni aussi d'excroissances sensorielles. Nereis dispose donc d'une série d'organes des sens beaucoup plus complexe que notre ver de terre. En effet, il est plus explorateur que le Lombric. Nereis possède aussi une paire de mâchoires cornées qui sont enfouies dans le corps. Pour se défendre, ou pour saisir une proie, l'animal dévagine son pharynx et sort ses deux mâchoires, les déployant largement. La morsure qu'infligent ces dents peut être douloureuse, surtout quand Nereis essaie ensuite de rétracter son pharynx. Si ce n'est pas le doigt résistant d'un pêcheur que Nereis a accroché, mais un Mollusque, un autre ver, un crustacé, il est capable ainsi d'en arracher un morceau et de l'engloutir en rétractant son pharynx.
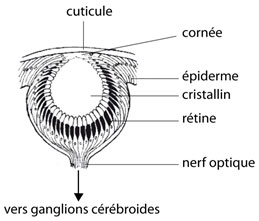
Oeil d'un polychète
Détaillons maintenant la structure d'un œil. Chacun des 4 yeux est constitué d'une cupule fermée. Le tégument, comme chez le Lombric, est un épithélium simple, recouvert d'une cuticule. Il recouvre l'œil et à cet endroit, il est transparent. Il reçoit le nom de cornée. La rétine est composée de cellules photoréceptrices, de cellules pigmentaires et de cellules de soutien. Les axones des cellules photoréceptrices se rassemblent en nerf optique. L'intérieur de la cupule oculaire est empli d'une gelée transparente, le cristallin, qui concentre les rayons lumineux sur la rétine.

Reproduction
Nous avons déjà évoqué la capacité de régénération des vers de terre. Celle des Polychètes est tout aussi grande, et même plus. Profitant en quelque sorte de leurs grandes capacités de régénération, de nombreux polychètes sont capables de se reproduire simplement par bourgeonnement. Par contre, les Achètes ont pratiquement perdu cette possibilité; cette perte probablement liée à la perte de la métamérisation typique, comme nous le verrons.
Les Polychètes errants présentent les sexes séparés : ils sont dioïques. Quelques sédentaires sont hermaphrodites, et l'hermaphrodisme est la règle chez les Oligochètes et les Achètes. L'apparition de l'hermaphrodisme peut s'expliquer par la faible probabilité de rencontre entre sexes : dès que deux hermaphrodites se rencontrent, l'autre est utilisable pour la copulation... mais si un mâle rencontre un mâle, il n'y a aucune chance que cette union soit fructueuse ! Donc dans un habitat aux rencontres aléatoires, la possession des deux sexes à la fois est un avantage pour la multiplication de l'espèce. Ce n'est pas la première fois que nous le constatons : pensez aux vers parasites Platyhelminthes.
Le développement chez les Polychètes passe par un stade larvaire libre et nageur: la larve trochophore que nous avons étudiée en détails dans la partie du cours consacrée au développement. Chez les Oligochètes et les Achètes qui en dérivent, l'œuf se développe dans un cocon formé par le clitellum. L'embryogenèse en est toute troublée : plus de larve trochophore, mais n'empêche, les étapes typiques du développement des Protostomiens sont conservées.
Une curiosité tout-à-fait remarquable chez les Polychètes: ils mettent à profit leurs capacités de régénération et de bourgeonnement pour élaborer des modes de reproduction sexuée extraordinaires.

Polychètes immatures et matures
A gauche, vous voyez un Polychète immature, et à droite, ce même Polychète à maturité sexuelle. Que s'est-il passé ? Le ver a subi de profondes modifications internes et externes. Les produits génitaux mûrissent dans la partie postérieure du corps, dont l'aspect change. Ce phénomène porte le nom d'épitoquie. Le ver de droite est devenu épitoque. Il abandonne sa vie au fond de la mer pour devenir pélagique, nager en haute mer. Cette excursion se termine par l'émission des produits sexuels soit mâles, soit femelles.
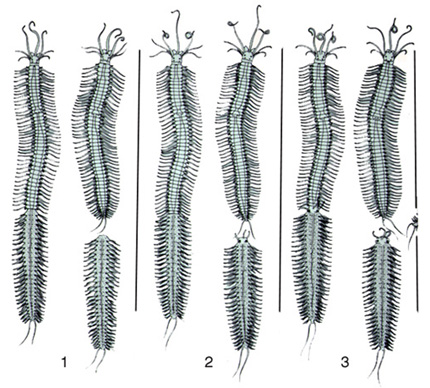
La scissiparité
L'épitoquie se combine parfois à la scissiparité. Ce phénomène est illustré ici en trois modalités différentes.
En 1, la souche, c'est-à-dire la partie antérieure de ce ver, a régénéré une région postérieure, tandis que le rejeton, qu'on appelle «stolon», demeure sans tête, acéphale.
En 2, le stolon, lui aussi, a régénéré une tête.
En 3, la régénération des parties manquantes se fait avant la scission du ver ; vous voyez que les yeux du stolon s'y trouvent déjà, avant même qu'il ne soit séparé de sa souche.

Les stolons
La performance de ce ver-ci est peu banale: c'est toute une chaîne de stolons bourgeonnants sexués qu’un individu forme.
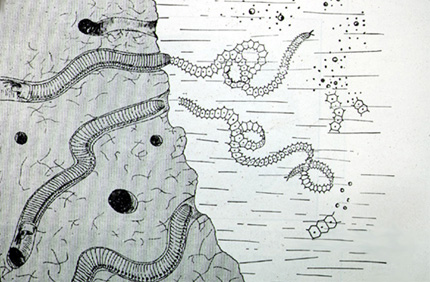
Eunice viridis
Voici Eunice viridis, un Polychète spectaculaire qui illustre encore cette stolonisation. Les polychètes du genre Eunice vivent dans les récifs coralliens des îles polynésiennes. Vous les voyez dans leurs galeries. La partie antérieure du ver, la souche, reste dans sa galerie alors que le stolon se détache pour mener une vie pélagique. Finalement, le stolon se fragmente et éclate pour libérer des ovules ou des spermatozoïdes. C'est un accouplement par procuration, les parents restent dans leurs galeries et régénèrent pour l'année suivante des nouvelles régions génitales. Mais cet accouplement de stolons se fait à un moment bien précis : les 7, 8 et 9èmes jours qui suivent la lune de novembre, les stolons bourrés de produits génitaux se détachent des souches et nagent tous en même temps. On les appelle «palolos». Les Palolos sont si nombreux qu'ils blanchissent la mer. Les indigènes polynésiens en sont très friands et ils préparent soigneusement la pêche et les fêtes qui l'accompagnent.

Nereis virens
Nous allons illustrer rapidement maintenant la diversité des Polychètes, riches d’environ 10000 espèces connues.
Nereis virens est une autre espèce du même genre que Nereis diversicolor, que nous connaissons déjà. Celle-ci est plus grande : une trentaine de centimètres. Elle nage aussi au niveau des fonds sableux marins et se creuse des galeries de passage.

Pterosyllis sp
Pterosyllis sp. mesure 10 à 15 millimètres. Ce Polychète errant a les parapodes garnis de longs filaments. Il nage en pleine eau ou rampe sur les algues.

Aphrodite
La métamérisation d’Aphrodite, la Souris de mer, est masquée par les longues soies qui garnissent sa face dorsale, mais elle est apparente sur sa face ventrale. Les Souris de mer peuvent atteindre une longueur de 15 centimètres, excédant de loin la taille des Mammifères dont elles portent le nom. Aphrodite vit dans la zone sublittorale, enfoncée dans le sable ; seule l'extrémité postérieure émerge.
Nereis, Peterosyllis, Aphrodite sont des Polychètes qualifiés d’errants : ils se déplacent librement dans l'eau et s'ils s'enfouissent dans des galeries, ce n'est que transitoire. Ils ne sont pas attachés à ce lieu. Ces vers sont généralement carnassiers, ils se nourrissent de proies vivantes plus ou moins volumineuses, mais ne dédaignent pas quelques plantes.

Les spirographe
Les Polychètes qualifiés de sédentaires vivent soit dans un tube qu'ils sécrètent, soit dans une galerie qu'ils consolident de mucus. Ces sédentaires se nourrissent habituellement de plancton en suspension dans l'eau, et de petites particules.
Le Spirographe que vous voyez sécrète un tube membraneux à consistance élastique dans lequel il vit. Seule la couronne de filaments branchiaux sort du tube. Grâce à ces filaments, le ver capte sa nourriture. Le Spirographe mesure une vingtaine de centimètres de long.

La serpule
Cette Serpule vit aussi dans un tube qu'elle sécrète. Ce tube est calcaire, très résistant et fixé au rocher.
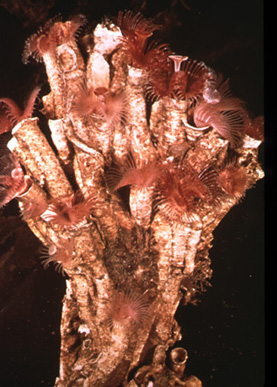
La serpicule
Voici une espèce de Serpule dont les tubes individuels s'accolent les uns aux autres, édifiant ainsi une sorte de récif. L'ouverture du tube peut être obturée par un petit opercule.

La serpicule
On voit ici un opercule, situé au sommet de l’organisme et de couleur blanche. Il est rigide et porté sur la tête. Quand le ver se retire brusquement dans son abri, l'opercule se place dans l'ouverture du tube, à la façon d'un bouchon dans une bouteille. On peut sortir les Serpules de leur tube, qui ne colle pas au tégument à la façon d’une carapace, bien que ce soit le tégument qui le sécrète.

La lanice
La Lanice s'abrite dans un étui formé de débris de coquillages agglutinés par une membrane très souple que sécrète le ver. A marée basse, les plages de nos régions sont hérissées de ces tubes. Ils sont très longs et profondément enfoncés dans le sable ; seule l'extrémité dépasse la surface de 4 à 5 centimètres. A marée basse, le ver se réfugie tout au fond de son long tube.

Arénicole
Vous avez déjà vu des plages couvertes de petits tortillons de sable. D'où proviennent ces tortillons ? A une petite distance de chaque tortillon, on trouve un orifice en entonnoir. Si on creuse très rapidement à ce niveau, on peut dénicher l'Arénicole.

Arénicole
L'Arénicole mesure 20 à 25 centimètres. Lorsqu'on le touche, il laisse suinter un liquide qui tache fortement les mains en jaune-orange. Des houppes de branchies rouge-vif garnissent les côtés des segments antérieurs, mais il n'en existe pas sur la partie postérieure du corps, plus grêle, qui ressemble à une queue. L'Arénicole possède un pharynx évaginable, et utilise cet outil pour fouir le sable humide. Quand il se nourrit, le ver avale en même temps, presque sans discrimination, les matériaux du fond, au rythme d'une bouchée toutes les cinq secondes environ. Il continue à cette allure pendant une minute, puis se repose deux à trois minutes, et ainsi de suite. Ainsi, le quart de ces journées est employé à se nourrir avec des ascensions occasionnelles à la surface du sable pour expulser un tortillon par l'anus.
Exercice
Résumez les adaptations évolutives chez les Polychètes errants et sédentaires.

Le ver de terre
L'exemple type d'Annélide que nous avons choisi, le Ver de terre, est placé au sein du clade des Oligochètes : par métamère, il dispose de 4 paires de soies.
Le Tubifex
Parmi les Oligochètes, on trouvera cependant aussi des espèces dulçaquicoles, telles le Tubifex.
Les Tubifex, rouges vifs ou brunâtres, prospèrent dans les fonds vaseux riches en matières organiques et peuvent se contenter de très peu d'oxygène ; ce sont d'ailleurs des indicateurs de pollution organique. Dans ces eaux, ils se rassemblent en une sorte de tapis animal.
Quand il n'est pas inquiété, le Tubifex grimpe à l'extrémité de son tube muqueux, qui forme une espèce de tunnel vertical dans la vase. Son corps long de 2-3 centimètres, ondule dans l'eau ; il avale la vase et la rejette à l'autre bout. Au moindre danger, chaque Tubifex se réfugie instantanément dans son tube.
Ceux parmi vous qui ont un aquarium doivent bien les connaître ; on les donne comme nourriture aux poissons.

Hirudo medicinalis
Les Achètes ou Hirudinées regroupent des Annélides sans soies. C'est le cas de la Sangsue Hirudo medicinalis. L'extrémité antérieure de cette Sangsue est modifiée en ventouse buccale tandis que l'extrémité postérieure forme une ventouse au-dessous de l'anus. La sangsue est très facile à manipuler ; elle se fixe d’elle-même à l'endroit qu'on lui assigne.
La Sangsue médicinale suce le sang, sa nourriture de choix. Grâce à un enzyme salivaire, l'hirudine, qui liquéfie les caillots, elle peut faire disparaître une meurtrissure trop visible, comme un «bleu» ou un «œil au beurre noir». Autrefois, les médecins, très portés sur les saignées, les utilisaient pour tirer peu à peu le «mauvais sang». Après la saignée, inutile de tirer sur la sangsue pour l'ôter ; elle est bien accrochée par ses petites dents. L'application d'un peu de sel sur la sangsue gorgée la fait se détacher et dégorger. On la lave à l'eau douce, et elle est à nouveau prête à un autre repas. Il n'y a pas si longtemps, on se la procurait encore en pharmacie...

Oligochètes d'eau douce
Toutes les Sangsues ne sucent pas le sang ! Nombres d'entre elles ne sont pas des ectoparasites, mais toutes possèdent des ventouses. Elles dériveraient d'Oligochètes d'eau douce, qui abandonnèrent le fouissage, pour exploiter la surface des fonds aquatiques. Elles nagent gracieusement en ondulant leur corps, mais à la fin de la journée, elles s'accrochent à un support solide par la ventouse située à l'extrémité postérieure du corps. Souvent, elles présentent une seconde ventouse antérieure.

Déplacement des sangsues
Les sangsues peuvent aussi ramper sur un support solide. La ventouse postérieure étant fixée, les muscles circulaires se contractent ; la sangsue s'allonge, attache sa ventouse antérieure. Puis la ventouse postérieure se détache et les muscles longitudinaux se contractent au maximum, de telle sorte que la ventouse postérieure soit ramenée vers l'avant, juste derrière l'antérieure. Ainsi, chaque «enjambée» est presqu'égale à la longueur de l'animal étiré.
Au contraire de la locomotion péristaltique du ver de terre, ce mode de reptation ne nécessite aucune différence de pression interne selon les segments : la musculature totale se comporte comme une unité. La fonction mécanique des dissépiments a disparu... ils disparaissent (ils ne feront qu'une brève apparition lors du développement embryonnaire). En plus, la sangsue se contente de ramper sans s'enterrer; elle n'a besoin que de forces réduites : le cœlome hydrostatique se réduit.
La métamérisation n'est plus évidente, mais garde des traces dans la structure des organes internes. Par contre, la segmentation externe visible sur le tégument n'y correspond pas ; il s’agit d’une annellation secondaire.
Comme les Oligochètes, les Achètes sont hermaphrodites et possèdent un clitellum, moins visible cependant. Les Achètes sont surtout dulçaquicoles; ils ont réalisé quelques colonisations du milieu terrestre, et ensuite quelques-uns sont retournés secondairement à la mer.
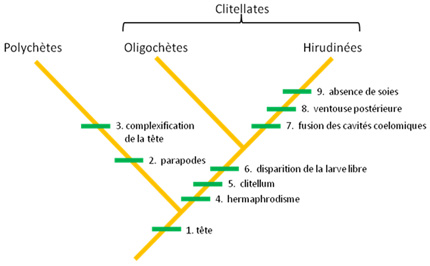
Evolution des Annélides
Ce cladogramme présente les principales étapes de l’évolution des Annélides. L’origine des Annélides aurait été marquée par l’apparition de (1) la tête formée de segments : prostomium et péristomium. Une différenciation de cet ancêtre commun à tous les Annélides a mené à la lignée des Polychètes (Polychaeta) caractérisée par l’apparition de parapodes (2) et la complexification de la tête (3).
Le clade qui mène à la lignée des Clitellates est défini par l’apparition des caractères suivants: (4) l’hermaphrodisme, (5) le clitellum épidermique, (6) la disparition de la larve libre.
Les oligochètes forment le groupe plus ancestral des clitellates, parce que la lignée des hirudinées est caractérisée par l’apparition des synapomorphies supplémentaires suivantes : (7) la fusion des cavités cœlomiques et la réduction du nombre de dissépiments, (8) l’apparition d’une ventouse postérieure, (9) la perte des soies sur le corps.