Les Cnidaires
Ils sont le premier grand Embranchement de Métazoaires au sens strict. Leurs aspects sont très variés (Hydre, Méduse, Anémone de mer, récifs de Coraux), mais peuvent se ramener à deux formes fondamentales : le polype et la méduse. L'habitat est aquatique, surtout marin, avec quelques familles en eau douce. Nous étudierons 4 représentants bien concrets, puis dégagerons les caractères essentiels de leur anatomie et de leur cycle vital, qui permettent de proposer une classification et un schéma évolutif.
Fiche récapitulative
- Premier grand embranchement de Métazoaires comportant de vrais feuillets embryonnaires (ectoderme, endoderme), ils sont diploblastiques ;
- 99% sont marins, 1% dulçaquicoles ;
- Embranchement très diversifié incluant les méduses, les anémones de mer, les coraux et les hydres ;
- Vivent fixés sur un support ou nagent librement ;
- Au stade adulte leur symétrie est radiaire, on retrouve chez les cnidaires le stade polype et le stade méduse ;
- Plus de 9000 espèces décrites ;
- L’origine date du Pré-cambrien ;
- Ils contiennent des cnidocytes étant des cellules spécialisés, propre à ce groupe, qui servent à la défense ;
- Une cavité gastro-vasculaire sert à la digestion et la circulation des éléments nutritifs ;
- Système nerveux : un réseau de cellules nerveuses ;
- Absence de système respiratoire et excréteur, cela se fait par diffusion ;
- Larve caractéristique des cnidaires: la planula ciliée, à symétrie bilatérale.
1. Présentation du groupe
Les Cnidaires sont généralement marins, cependant, quelques familles vivent en eau douce.
L'embranchement des Cnidaires est très prolifique. Dans certaines mers chaudes, ils occupent le moindre centimètre carré du fond, dominant la vie des autres non vertébrés et même des poissons.
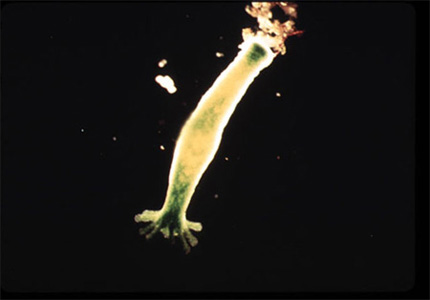
A première vue, cet embranchement est constitué d'animaux très dissemblables. Sur cette image, vous voyez une hydre d'eau douce. Sa couleur verte est donnée par une symbiose avec des algues photosynthétiques se trouvant à l’intérieur des cellules de l’endoderme. Des algues vertes parasites se trouvant à la surface de l’hydre peuvent également augmenter la coloration verte.
Les méduses et les anémones de mer appartiennent au même groupe, tout comme les coraux.

Une méduse

Une anémone de mer
Fixée sur un mollusque gastéropode.
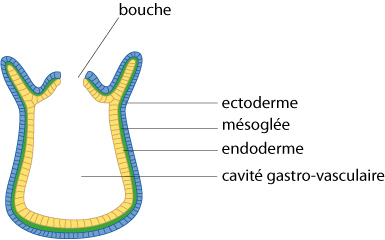
La cavité gastro-vasculaire
Pourtant en examinant plus attentivement ces animaux, on découvre qu'ils sont tous bâtis selon une structure fondamentale commune. Cette structure est schématisée sur le dessin. Leur corps est formé d’une urne, possédant une seule ouverture, la bouche, et une vaste cavité interne. Cette cavité est appelée cavité gastro-vasculaire, parce qu'elle sert à la fois à la digestion et à la circulation des éléments nutritifs. La présence d'une bouche et d'une grande cavité digestive permet à ces animaux de se nourrir de proies plus grandes que ne peuvent les Spongiaires. Les Cnidaires sont des macrophages : ils se nourrissent d’organismes non vertébrés divers, et même de petits poissons.
La paroi de leur corps est constituée d'ectoderme, à l’extérieur, et d'endoderme, à l’intérieur, séparés par une couche de mésoglée, une matrice essentiellement acellulaire qui peut être colonisée par quelques cellules issus des feuillets cellulaires.
Par convention, on représente l'ectoderme en bleu, l'endoderme en jaune. La mésoglée est représentée en vert et plus tard, les tissus dérivés du mésoderme, seront en rouge.
Un mollusque gastéropode
Comme nous l'avons vu sur les photos, cette structure commune aux Cnidaires s'exprime sous diverses formes. Certains Cnidaires vivent fixés sur un support, d'autres au contraire nagent librement dans l'eau. Regardez la photo de cette anémone de mer . Elle illustre la forme fixée. Vous y trouvez la structure d'une urne allongée, avec la bouche entourée d'une couronne de tentacules à une extrémité. A l'autre extrémité, qualifiée d’extrémité aborale, parce que située du côté opposé à la bouche, se trouve le pied qui adhère au support, dans ce cas-ci, un mollusque gastéropode.
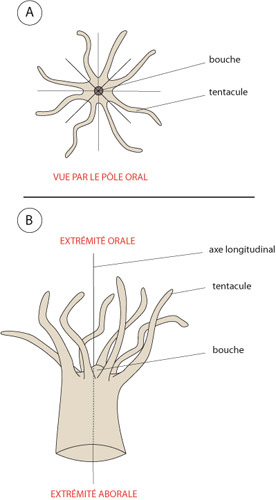
La symétrie radiaire
L'axe longitudinal allant de la bouche au pied s'appelle l'axe oral-aboral. Le corps de l'individu présente une symétrie radiaire par rapport à cet axe. La figure vous le montre en schéma. Tout Cnidaire ayant un mode de vie fixé (on dit qu’ils sont sessiles), porte le nom général de polype.
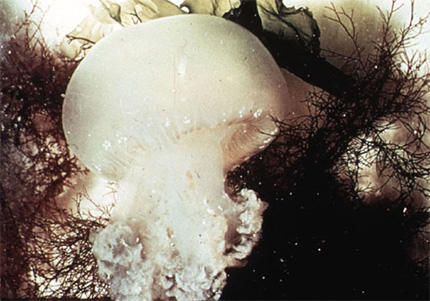
La méduse
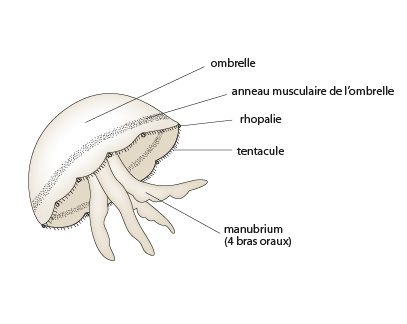
Cette photo vous montre une méduse. Tout Cnidaire ayant un mode de vie libre dans l'eau porte le nom général de méduse. La bouche, prolongée par une sorte de trompe, le manubrium, est tournée vers le bas. Le pôle aboral est convexe et forme une coupole, l'ombrelle. La symétrie est radiaire suivant l'axe oral-aboral tout comme chez le polype.
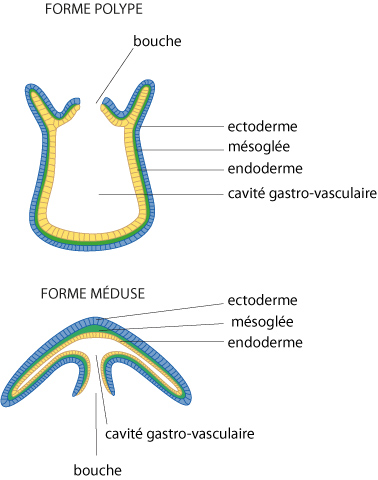
Structure des polypes et des méduses
Cette figure illustre la similitude de structure des polypes et des méduses : forme d'urne pour le polype, forme d'urne renversée et surbaissée pour la méduse; une seule ouverture, la bouche; deux feuillets cellulaires séparés par une couche de mésoglée. Cette couche est nettement plus épaisse au pôle aboral des méduses, formant ainsi l'ombrelle.
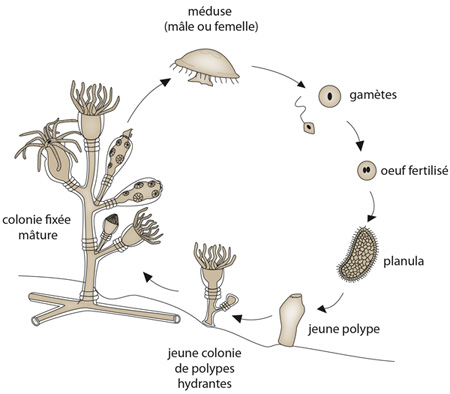
Cycle de vie des cnidaires
Nous allons examiner maintenant le cycle de vie des Cnidaires. Certaines espèces ne comportent que le stade polype, d’autres uniquement le stade méduse. Certaines espèces vont alterner les deux formes au cours de leur cycle vital. Regardez le polype à gauche de l'image. D'un de ses rameaux, émerge une petite méduse qui est propulsée dans le milieu aquatique. Lorsqu'elle sera adulte, elle produira des spermatozoïdes si elle est mâle, des ovules si elle est femelle. La fécondation d'un ovule par un spermatozoïde donne un œuf, un zygote qui se transforme en larve. La larve se fixe au sol et se développe en polype; le cycle polype-méduse continue de génération en génération.
Les Cnidaires vivent soit en colonies, soit sont solitaires, selon les espèces ou l’étape de leur cycle. Sur cette image, vous voyez à gauche une colonie de polypes. Par contre, la méduse a un mode de vie solitaire.

2. Exemples-types - 2.1. L'anémone de mer Actinia equina (Anthozoa, Hexacorallia)
Maintenant nous allons passer à une étude plus approfondie des Cnidaires. Dans ce but, nous ne les considérerons plus en bloc comme un embranchement, mais nous évoquerons des animaux bien réels qui vivent dans les mers ou les eaux douces.
Les Anémones de mer appartiennent aussi à l'embranchement des Cnidaires. Le nom du genre de celle-ci est Actinia. Vous en voyez une ici, fixée sur un rocher. Sur les côtes rocheuses de la Manche, on peut les apercevoir à marée descendante. Mais c'est dans les mers chaudes qu'on les trouve en abondance, ainsi que de nombreuses espèces voisines souvent brillamment colorées. Les Anémones ne forment pas de colonies, chaque individu mène une vie solitaire. Par ailleurs, le cycle de vie ne contient que le stade polype: il n’y a pas de stade méduse chez cet organisme.

2.1.1. Examen externe
Le tronc de l’anémone de mer est généralement court. La base du tronc, soit le pôle aboral, lui sert de ventouse par laquelle elle se fixe au support. Remarque, a marée basse, l’anémone replie ses tentacules afin de limiter l’évaporation de l’eau et contracte son tronc afin de réduire sa surface. En travaux pratiques, vous verrez nettement ce disque musculeux grâce auquel elle peut glisser et ainsi se déplacer lentement. Elle peut aussi se détacher complètement de son support et nager maladroitement en se tortillant.

Pôle oral de l'Anémone
Voici le pôle oral de l'Anémone. Au centre se trouve la bouche en fente allongée. Plusieurs cercles de tentacules l'entourent. L'aspect de fleur de l'Anémone de mer lui a valu le nom de polype anthante du grec « anthos », fleur.
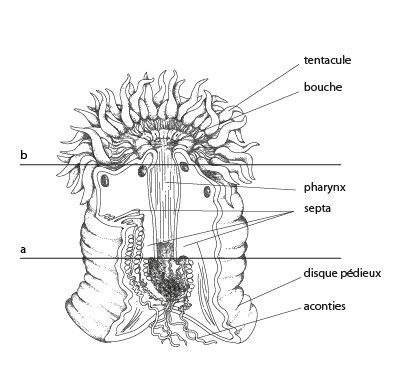
2.1.2. Examen interne - 2.1.2.1. Tégument
Voici une coupe longitudinale d'Actinie. La bouche est prolongée d'un pharynx musculeux. Ce pharynx descend jusqu’au tiers environ de la cavité gastro-vasculaire. On pourrait le comparer au tube d'un entonnoir.
On voit aussi des cloisons verticales dans la cavité gastro-vasculaire (côté aboral): ce sont les septa.
Coupe transversale du polype
En « a » nous avons schématisé une coupe transversale du polype. Cette coupe est située au niveau a de la figure précédente. Les septa sont des replis de l'endoderme ; ils peuvent être fort nombreux. Chez une jeune anémone, on en compte 6 paires. Le schéma « b » est une coupe transversale au niveau du pharynx. Donc, ces replis de l'endoderme forment des logettes, fermées au niveau du pharynx et ouvertes plus bas. L'endoderme des septa et celui du pharynx, en jaune sur la dia, sont en continuité.
Coupe transversale dans le tronc d'Actinie
Cette photo montre une coupe transversale sous le pharynx. Les septa sont libres dans la cavité gastro-vasculaire. On voit que la réalité n'est jamais aussi rigoureusement géométrique qu'un schéma. Sur chacun des septa, les épaississements sont les fibres musculaires de l'endoderme.
Les replis de l'endoderme sont appelés septa mésentériques. Les logettes sont aussi qualifiées de mésentériques.
Nous avons dit qu'une jeune Anémone possède 6 paires de septa mésentériques. Au cours de sa croissance, elle en acquiert de nouvelles paires; ce sont les septa secondaires.
Septa secondaires
La figure montre comment ces septa secondaires viennent s'intercaler entre les paires existantes. C'est un accroissement en cercle, qualifié de cyclomérique. Cet accroissement cyclomérique est continu. L'Anémone jeune à 6 paires de septa, puis 12, puis 24 et ainsi de suite. La disposition des septa dans la cavité gastro-vasculaire définissent les deux clades au sein des Anthozoaires (voir partie Diversité du groupe) et reflète également une symétrie bilatérale émergente.
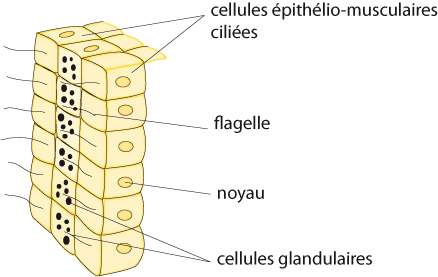
Cavité gastro-vasculaire
On peut voir aussi, sur le bord libre des septa, des rangées de cellules épithélio-musculaires flagellées entourant une rangée de cellules glandulaires. Ce regroupement de cellules identiques à des endroits déterminés constitue une ébauche d'organes. Ce fait marque une progression dans l'organisation cellulaire par rapport aux espèces animales que nous avons vues jusqu'ici. Cette structuration de l’ectoderme et de l'endoderme est semblable à celle qu’on retrouve chez les autres Cnidaires mais la disposition cellulaire de ces tissus est plus régulière chez cet anthozoaire. L’architecture de la cavité gastro-vasculaire ne modifie pas le processus de digestion dans ses grandes lignes. On a toujours d'abord une digestion extra-cellulaire et ensuite une digestion intracellulaire.
Exercice : citez quelles pourraient être les conséquences adaptatives de la complexification de la cavité gastro-vasculaire chez les Anthozoaires ?
2.1.2.2. Système locomoteur
Sera examiné chez la méduse Aurelia aurita.
2.1.2.3. Système digestif
Sera examiné plus en détail chez les hydrozoaires.
2.1.2.4. Système respiratoire
Il n’y a pas de systèmes respiratoire ni excréteur chez les Cnidaires. Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion: la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
2.1.2.5. Système circulatoire
Le système circulatoire est couplé au système digestif, comme nous le verrons chez les hydrozoaires.
2.1.2.6. Système excréteur
Il n’y a pas de systèmes respiratoire ni excréteur chez les Cnidaires. Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion : la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
2.1.2.7. Système nerveux
Sera examiné chez la méduse Aurelia aurita.
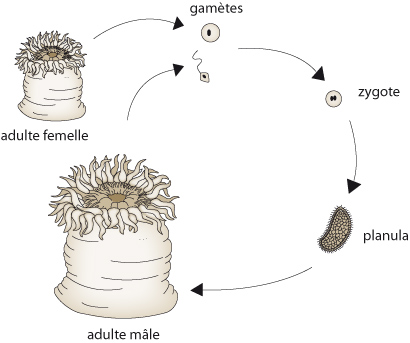
2.1.2.8. Système reproducteur et cycle de vie
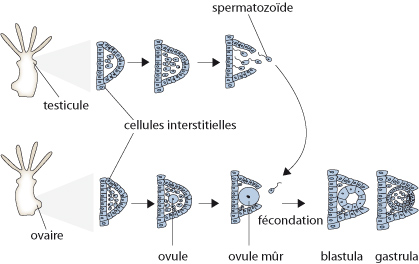
L'Anémone de mer est dioïque, c'est-à-dire qu'elle est soit mâle, soit femelle. La figure schématise son cycle vital. Les ovules et les spermatozoïdes se développent dans les septa. Ils sont expulsés de la cavité gastro-vasculaire par la bouche et la fécondation se produit dans l'eau. Le zygote se développe en une larve planula libre à symétrie bilatérale qui se fixera dans une crevasse rocheuse et formera une anémone adulte.
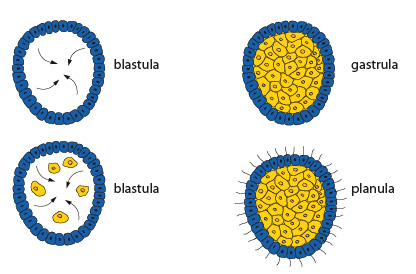
Les stades de l'embryon
Les stades de l'embryon sont ici, schématisés en coupe. D'abord, le stade blastula. C'est le stade précoce d'un embryon dont les cellules déjà nombreuses forment une sphère creuse. La cavité se nomme blastocœle. Par la gastrulation, la blastula se transforme en une gastrula à deux feuillets cellulaires: l'endoderme se forme par migration de certaines cellules périphériques dans le blastocœle, comme les flèches l'indiquent. Le blastocœle se remplit complètement de ces cellules endodermiques, jusqu’à disparition complète de la cavité. Les cellules du feuillet extérieur de la gastrula acquièrent des cils qui permettront à la larve de nager. A ce moment, on l'appelle planula. Cette information est importante parce que vous retrouverez ces stades dans les groupes taxonomiques plus évolués.
L'Anémone peut parfois se reproduire d'une façon asexuée par scission; ce phénomène s'appelle la scissiparité. Elle se fend longitudinalement en deux et chaque moitié reconstruit la moitié manquante. Bien sûr, ce processus se fait très progressivement. Une reproduction asexuée par bourgeonnement survient aussi quelques fois. Le nouveau bourgeon se détache de l’animal-mère, formant un nouvel individu. Il n'y a jamais formation de colonies stables, telles que le font des cnidaires qui leur sont proches, les coraux.
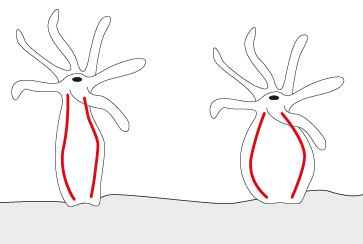
Les grandes Anémones de mer vertes
Les grandes Anémones de mer vertes vivent dans les eaux froides de côtes rocheuses. Les deux individus que l’on voit mesurent environ 25 cm de diamètre et auraient l’âge de 15 ans. Tout poisson ou crustacé qui effleure les tentacules urticants est paralysé et englouti par la bouche. Certaines Anémones bourgeonnent des individus-fils à la surface de leur corps. Ces rejetons se détacheront pour trouver un autre habitat.
Les Coraux ont la même structure générale que l’Anémone, mais ils secrètent un squelette. Ils restent attachés en colonies de centaines d’individus, aux formes très variées.

2.2. La méduse Aurelia aurita (Scyphozoa) - 2.2.1. Examen externe
Voici un autre Cnidaire. C'est Aurelia sp., une méduse commune en Europe. Bien qu’une forme polype existe pour cette espèce, on la retrouve surtout sous sa forme méduse. Elle mesure de 10 à 40 cm de diamètre. Sous l'ombrelle, vous voyez 4 sortes de fer à cheval opaques: ce sont les gonades. Les méduses portant un voile portent le nom de méduse « craspédotes ».
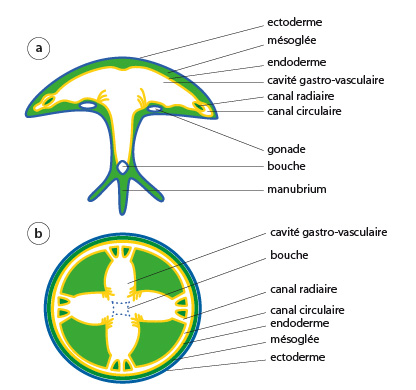
Coupes dans la cavité gastro-vasculaire de la méduse
En « a » se trouve un schéma d’une méduse vue en coupe longitudinale, en « b » en coupe transversale (ou vue ventrale).
Aurelia possède la structure fondamentale propre aux Cnidaires, c'est-à-dire un ectoderme et un endoderme. La cavité gastro-vasculaire, dont vous pouvez voir deux coupes ici, a une architecture plus complexe que celle des polypes. Regardez le schéma « b ». La bouche mène dans une large cavité gastro-vasculaire divisée en 4 lobes. Chaque lobe est prolongé par des canaux radiaires qui se dirigent vers la périphérie de l'ombrelle pour aboutir au canal circulaire. Au schéma « a », vous retrouverez ces éléments : la bouche, la cavité lobée, les canaux radiaires et le canal circulaire. La digestion s'opère en deux temps : extracellulaire d'abord, puis intracellulaire.
2.2.2. Examen interne
2.2.2.1. Tégument
Le tégument externe est formé par l’ectoderme, dessous se trouve la mésoglée, comme chez l’anémone.
2.2.2.2. Système locomoteur
La méduse se déplace constamment, aussi, elle a besoin de muscles mieux développés que les polypes. Les extensions des cellules épithélio-musculaires contenant les myofibrilles sont particulièrement bien développées sur le pourtour de l'ombrelle. La figure montre l'anneau musculaire puissant au bord inférieur de l'ombrelle. Une forte contraction de cet anneau provoque une éjection brutale de l'eau contenue dans l'espace sous-ombrellaire. Par effet de recul, l'animal se déplace.
2.2.2.3. Système digestif
La cavité gastro-vasculaire est quadrilobée. Chaque lobe est prolongé vers la périphérie par des canaux radiaires, qui se jettent dans un canal circulaire. La digestion s'opère en deux temps : extracellulaire, puis intracellulaire (en verra cela plus en détail chez les hydrozoaires).
2.2.2.4. Système respiratoire
Il n’y a pas de systèmes respiratoire ni excréteur chez les Cnidaires. Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion : la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
2.2.2.5. Système circulatoire
Il est couplé au système digestif chez les Cnidaires, comme nous le verrons chez l’hydre d’eau douce.
2.2.2.6. Système excréteur
Il n’y a pas de systèmes respiratoire ni excréteur chez les Cnidaires. Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion : la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
2.2.2.7. Système nerveux
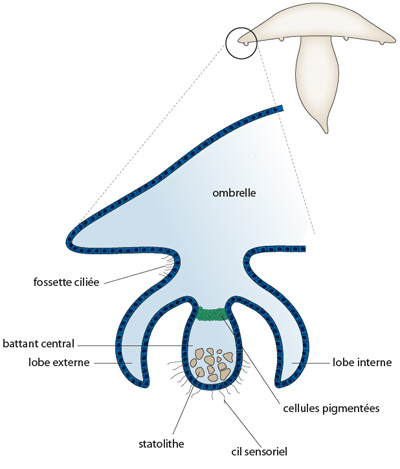
Les méduses possèdent des organes des sens sur le rebord de l'ombrelle. C'est la première fois que nous employons le terme d'organe. Ces organes portent le nom de rhopalies. Le schéma du bas situe les rhopalies. La fenêtre indique la partie d'ombrelle et la rhopalie qui sont schématisées en grand format et en coupe longitudinale. Le battant central en forme de massue est bourré de granulations calcaires, de sorte que son poids le maintient toujours plus ou moins vertical. Il est muni de cils sensoriels. Lorsque la méduse nage, ce battant est heurté par l'un ou l'autre des lobes qui l'entourent, soit le lobe interne, soit le lobe externe selon l'inclinaison que prend la méduse. L'animal est ainsi prévenu, en quelque sorte, de la position dans laquelle il se trouve. La rhopalie est donc un organe de l'équilibre. Le battant central porte aussi à sa base des cellules pigmentées qui semblent être sensibles à la lumière. La rhopalie comporte encore des fossettes ciliées qui paraissent activées par des composés chimiques présents dans l’eau de mer. Ce sont des organes olfactifs.
Des cellules nerveuses sont annexées à la rhopalie. Le réseau nerveux coordonne donc la perception des stimuli extérieurs et la réponse adaptative de l'animal.
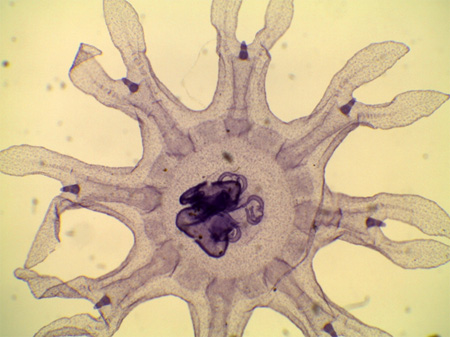
2.2.2.8. Système reproducteur
Les sexes sont séparés, mais les méduses mâles sont extérieurement semblables aux méduses femelles. La méduse possède 4 gonades situées sous l'ectoderme de la sous-ombrelle. A maturité, les ovules ou les spermatozoïdes percent l'endoderme, et sont libérés dans la cavité gastro-vasculaire puis expulsés dans le milieu extérieur par la bouche.
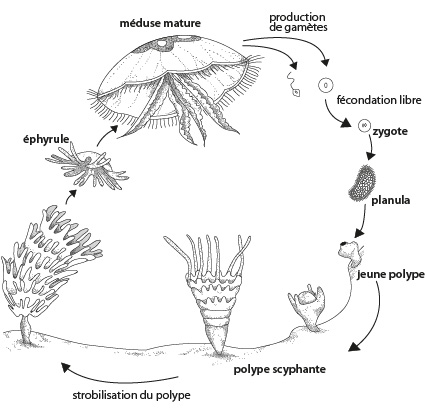
Cycle reproductif d'Aurelia Aurita
Cette figure se trouve aussi dans le syllabus. La fécondation est libre dans l'eau. Le zygote se segmente, donne une blastula, puis la gastrulation spéciale des Cnidaires forme une larve planula ciliée. Elle comprend les deux assises cellulaires typiques des Cnidaires, l'ectoderme et l'endoderme. Elle se fixe au sol et forme un jeune polype. Une bouche apparaît et s'entoure de tentacules. Ce polype très vorace en forme de coupe est appelé polype scyphante, du grec scyphos, coupe.
Après que le polype a atteint une certaine taille, il se modifie par une série d'étranglements transversaux. Ce phénomène s'appelle la strobilisation. Il prend alors l'allure d'une pile d'assiettes superposées. Les segments s'étranglent de plus en plus, se détachent l'un après l'autre en commençant par le haut. Les petits disques flottants de 2 ou 3 millimètres de diamètre, appelés éphyrules, s'accroissent pour devenir des méduses adultes qui produiront de nouveaux polypes par voie sexuée. Chez Aurelia, il y a donc alternance d'une forme polype de petite taille et de la forme bien connue, de grande taille, la méduse.

2.3. La colonie d'Obelia geniculata (Hydrozoaire) - 2.3.1. Examen externe
Voici une colonie de polypes d’Obelia, qui vit en eau de mer. On peut facilement en trouver sur les côtes de la mer du Nord aux endroits rocheux. Obelia se présente en petite colonie buissonneuse de 3 à 5 cm de haut. Elle est attachée aux rochers ou à des coquilles, le long des côtes à faible profondeur. De petits bourgeons se forment. Ce sont des nouveaux individus. Ils restent attachés au tronc mère (le stolon) et la colonie grandit.
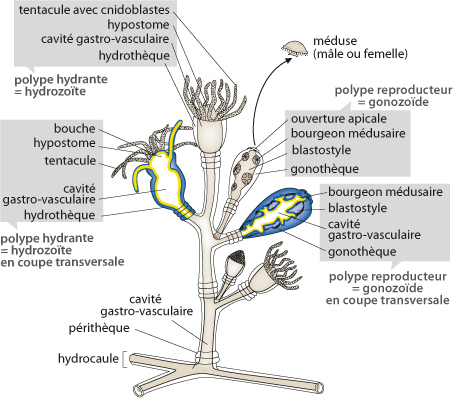
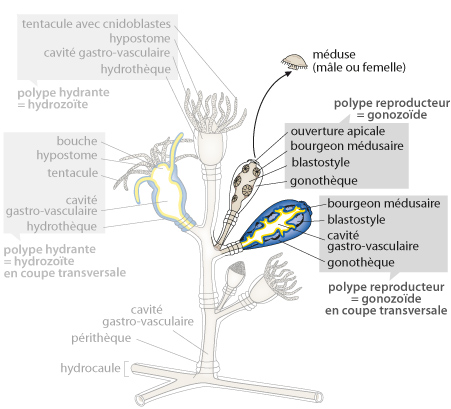
Obelia Geniculata
Cette figure, que vous trouvez aussi dans le syllabus, reproduit d'Obelia. Les polypes dont le pôle oral est entouré de tentacules sont appelés polypes hydrantes. Au centre du schéma, vous en voyez un dessiné en vue réelle, à gauche, il est dessiné en coupe. A droite, un polype hydrante aux tentacules rétractés. Ces tentacules ont pour rôle d'émettre un venin qui paralyse les proies, puis de les acheminer vers la bouche.
Le schéma vous montre d'autres polypes encore. Ils n'ont pas de tentacules, ce sont les polypes reproducteurs appelés aussi gonozoïdes. A droite de la figure, vous voyez un polype reproducteur à l'état naturel, à côté il est en coupe. Le rôle du gonozoïde est de bourgeonner des petites méduses qui abandonneront le polype reproducteur. La colonie toute entière est protégée par une gaine de chitine sécrétée par l'ectoderme. La chitine est un polymère organique résistant.
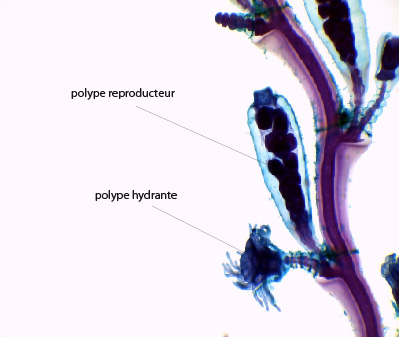
Obelia Geniculata
Voici une portion en photo d’une colonie d’Obelia geniculata.
Polype scyphante d’Aurelia
La différence essentielle entre un polype scyphante d’Aurelia et un polype hydrante d'Obelia ne réside pourtant pas dans cette forme extérieure mais dans la configuration de l'endoderme. Ce schéma montre en effet que l'endoderme du polype scyphante fait 4 replis, ce qui donne à la cavité gastro-vasculaire un aspect quadrilobé. Grâce à ces replis, la surface de contact entre l'endoderme et le milieu extérieur est fortement augmentée.
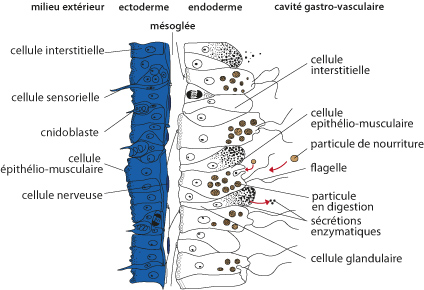
2.3.2. Examen interne - 2.3.2.1. Tégument
Passons à l'examen interne de l'Obelia. Cette figure, que vous trouvez aussi dans le syllabus, représente schématiquement une coupe longitudinale d'un polype hydrante. Vous repérez à gauche l'ectoderme, au centre la fine couche de mésoglée et à droite l'endoderme. L’organisation de ce tégument est similaire pour la plupart des Cnidaires ; nous allons le détailler chez cette hydre.
Examinons d'abord les cellules de l'ectoderme.
Il y a de nombreuses cellules appelées épithélio-musculaires. Dans les régions élaboratrices de la gaine de chitine, ces cellules secrètent les substances par leur pôle apical. La base de ces cellules est en contact avec la mésoglée et s'étale suivant l'axe oral-aboral de l'animal. Cette région contient un petit faisceau de fibrilles musculaires. Ces extensions sont en connexion les unes avec les autres, si bien que l'ectoderme forme dans son ensemble un manchon musculaire à fibrilles longitudinales.
Fibrilles musculaires
Quand les fibrilles longitudinales se contractent, le polype raccourcit. Vous en voyez une illustration sur la figure. A gauche, elles sont décontractées, à droite elles sont contractées.
Cellules sensorielles
Dans l'ectoderme, on trouve aussi des cellules sensorielles. Elles sont longues, minces, étirées et portent un cil à leur extrémité. Le cil, pointe avancée dans le monde extérieur, y capte les informations. La base de ces cellules se ramifie dans la mésoglée. Nous verrons tout à l'heure qu'elles sont en rapport avec les cellules nerveuses.
Toujours dans l'ectoderme, vous voyez les cnidoblastes. Ce sont des cellules caractéristiques des Cnidaires. Les cnidoblastes sont des cellules urticantes qui servent à la défense de l'organisme et à la capture des proies. Chaque cnidoblaste possède en plus du noyau et du cytoplasme, un cil sensoriel et une grande vacuole contenant le liquide urticant (l’actinocongestine) et paralysant. On trouve des cnidoblastes dans tout l'ectoderme de la colonie d'Obelia mais ils sont surtout concentrés sur les tentacules où ils sont rassemblés en boutons urticants.
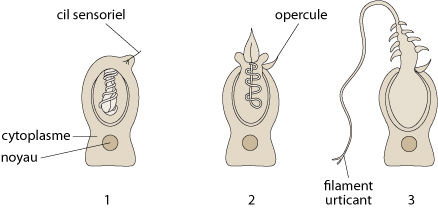
Fonctionnement des cnidoblastes
Voici comment le Cnidaire utilise ses cnidoblastes. Regardez le schéma 1, l'état de repos. La vacuole urticante est obturée par un opercule. A ce niveau, la paroi interne de la vacuole s'invagine en un tube creux enroulé sur lui-même. Cette paroi interne est armée de piquants. Lorsqu'une proie heurte le cil sensoriel, schéma 2, tout le cytoplasme se contracte. Sous la pression, l'opercule s'ouvre et le filament urticant commence à se dévaginer comme un doigt de gant. Schéma 3: les piquants du filament, maintenant sur la face externe vont s'enfoncer dans la proie et y produire de minuscules déchirures. Ces blessures sont suffisantes pour que le liquide de la vacuole qui s'écoule par l'orifice terminal du filament pénètre dans la proie et la paralyse ou la tue. Les tentacules n'ont plus alors qu'à ramener cette proie vers la bouche. Le cnidoblaste ne peut servir qu'une fois, après quoi, il doit être remplacé.
Cellules nerveuses
A la base de l'ectoderme, on trouve des cellules nerveuses, (bipolaires, tripolaires ou multipolaires). Ces cellules sont en relation les unes avec les autres au moyen de synapses. Elles innervent les cellules sensorielles, les fibrilles musculaires et les cnidoblastes. Il y a dès lors une connexion entre la sensation que reçoit le polype au contact d'un ennemi ou d'une proie et ses réactions musculaires ou de décharges des cnidoblastes.
Dans l'ectoderme encore, entre les différentes cellules spécialisées, nous voyons des massifs de cellules embryonnaires, non différenciées appelées cellules interstitielles. Elles peuvent se différencier en tous les types de cellules spécialisées que nous venons de voir.
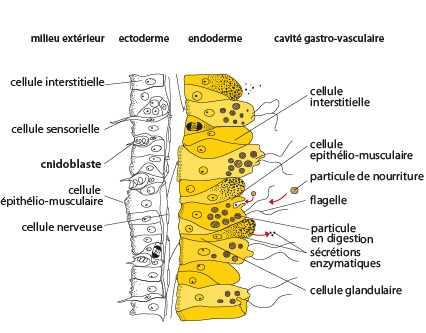
Composition cellulaire de l'endoderme
Voyons maintenant la composition cellulaire de l'endoderme.
Comme l'ectoderme, l'endoderme est surtout constitué de cellules épithélio-musculaires, mais ici, elles sont d'une taille supérieure et portent un flagelle. La base de ces cellules, c'est-à-dire la face en contact avec la mésoglée, s'étale transversalement dans la mésoglée en un réseau de fibrilles musculaires. Ces fibrilles sont représentées par les petits ronds à la base de la cellule. Etant transversales, elles forment une ceinture circulaire autour du polype.
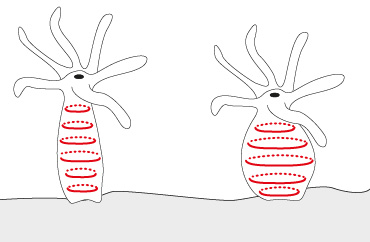
Contraction des fibres musculaires
Vous le voyez ici. La contraction de ces fibres provoque un allongement du polype. Les fibrilles musculaires de l'ectoderme et celles de l'endoderme travaillent donc dans un sens opposé. Ces cellules interviennent dans la contraction du polype, mais aussi dans la digestion.
Polype hydrante
Les flagelles des cellules épithélio-musculaires de l’endoderme, par leur battement, font circuler la masse d'eau présente dans la cavité gastro-vasculaire, et propulsent les particules alimentaires vers les parois. Les cellules épithélio-musculaires capturent ces particules par phagocytose, et les digèrent.
L'endoderme comporte aussi des cellules glandulaires. Elles déversent leur sécrétion dans la cavité gastro-vasculaire, ce qui permet la décomposition de la proie. En effet, les Cnidaires ne se contentent plus de particules nutritives minuscules comme les éponges, ce sont de macrophages. La décomposition de la proie par les enzymes digestifs dans la cavité gastro-vasculaire est une digestion extracellulaire. Elle se complète par la digestion intracellulaire qui se produit dans les cellules épithélio-musculaires.
Les autres types cellulaires de l'endoderme sont semblables à ceux de l'ectoderme. On ne trouve toutefois pas de cnidoblastes dans l'endoderme et les cellules interstitielles y sont plus rares.
Nous avons donc examiné les différents types de cellules que possède Obelia. La remarque suivante est importante: ces cellules ne sont pas condamnées à un emplacement fixe. De fréquents échanges de cellules se produisent entre l'ectoderme et l'endoderme à travers la mésoglée.
2.3.2.2. Système locomoteur
Les hydres ne se déplacent généralement pas. Elles sont fixées à leur substrat par la « sole pédieuse ».
2.3.2.3. Système digestif
La cavité gastro-vasculaire de chaque polype est en communication avec celle des autres. On peut dire que chaque individu est bien défini, que certains ont un rôle de capture et de digestion des aliments, d'autres ont un rôle reproducteur. Notons que les cavités des divers polypes communiquent via celle de la tige qui les porte. Cette cavité est donc digestive et vasculaire, ce qui justifie son nom : « gastro-vasculaire ». La nourriture captée par les polypes hydrantes est ainsi distribuée à toute la colonie. Voyez la section « Tégument » pour de plus amples informations sur la digestion chez l’hydre d’eau douce.
2.3.2.4. Système respiratoire
Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion : la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
2.3.2.5. Système circulatoire
Voyez les informations dans la section « système digestif ».
2.3.2.6. Système excréteur
Chez tous les Cnidaires, la respiration et l’excrétion se font par diffusion : la minceur de la paroi du corps et la circulation dans la cavité gastro-vasculaire permettent des échanges directs de toutes les cellules avec le milieu aquatique extérieur.
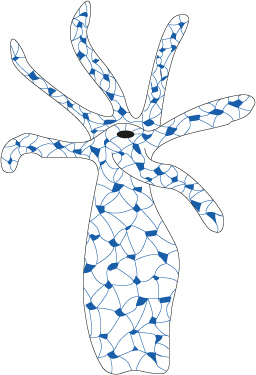
2.3.2.7. Système nerveux
Le réseau de cellules nerveuses constitue une première ébauche de système nerveux. C'est un système primitif, dépourvu de coordination centrale, dans lequel les influx circulent de manière multidirectionnelle. Vous pouvez voir ici une représentation simplifiée du réseau nerveux dans son ensemble. Ce réseau est plus dense sur la face orale du polype.
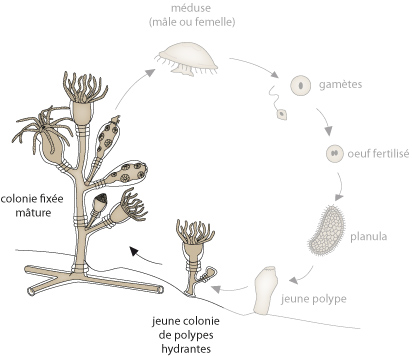
2.3.2.8. Système reproducteur
Examinons maintenant la reproduction d'Obelia. Elle se reproduit de façon asexuée par bourgeonnement comme vous voyez pour la jeune colonie formée. Le bourgeon s'accroît et non loin de son sommet des tentacules poussent, puis la bouche se forme. Un nouveau polype hydrante est constitué. Il reste attaché à la tige mère, ce qui accroît le nombre d'individus de la colonie.
Portion d'une colonie
Lorsque la colonie est bien développée, un autre type de polype apparaît par bourgeonnement également. Il s'agit des polypes reproducteurs ou gonozoïdes. On voit deux gonozoïdes ici. Ces polypes ne peuvent capter eux-mêmes leurs proies, les substances alimentaires leur parviennent par la cavité gastro-vasculaire commune.
Quelle est la structure de ce polype ?
Regardez sur la figure, le polype reproducteur de droite, il est vu en coupe. Il est constitué d'un axe creux fait d'ectoderme, tout comme la tige dont il a bourgeonné. Une gaine chitineuse l'entoure. Sur cet axe creux, des protubérances en forme de soucoupe apparaissent. Ce sont des bourgeons médusaires formés eux aussi des deux feuillets cellulaires et de mésoglée. Les bourgeons médusaires se détachent de l'axe et sont éjectés par une ouverture au sommet du gonozoïde. A ce stade, les petites méduses d'Obelia sont appelées gonophores. La figure vous donne un aperçu du phénomène. Les gonophores ont des tentacules sur le bord de l'ombrelle. Ils servent à capter les aliments. Au centre de la méduse, la bouche est prolongée en une sorte de tube, le manubrium. La bouche communique avec la cavité gastro-vasculaire.
Nous avons donc vu : le bourgeonnement de polypes hydrantes et le bourgeonnement de gonozoïdes.
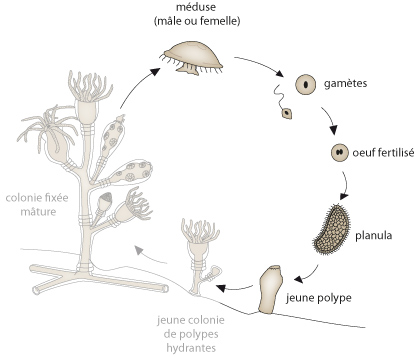
Voyons maintenant la reproduction sexuée d'Obelia
Les méduses produites, mâles ou femelles, possèdent quatre gonades situées sous l'ectoderme de l'ombrelle. Les produits génitaux doivent percer l'endoderme pour être évacués par la bouche. La fécondation de l'ovule se fait au hasard d'une rencontre avec un spermatozoïde dans l'eau, et le reste du développement suit celui vu pour l’anémone.
Après une série de divisions cellulaires, la planula se fixe au sol et perd ses cils. Une cavité se creuse dans le massif endodermique pour former la cavité gastro-vasculaire. Une bouche et des tentacules apparaissent, le polype fondateur d'une nouvelle colonie est constitué.
Exercice
Pour récapituler ce qui vient d'être dit, nommez et indiquez la fonction de tous les types d'individus qui existent au cours du cycle vital d'Obelia.
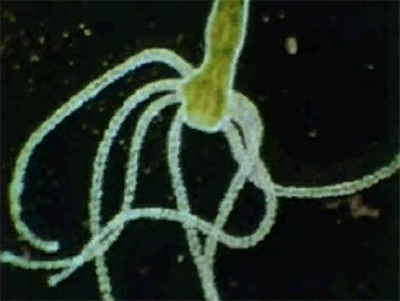
Les colonies d’Obelia
Les colonies d’Obelia se trouvent notamment sur les algues littorales. Elles sont composées de deux types d’individus : les polypes hydrantes et les polypes reproducteurs. Les polypes hydrantes capturent et digèrent la nourriture, qui est alors distribuée à la colonie, en passant par la cavité gastro-vasculaire commune. Cette cavité communique avec celle des polypes reproducteurs. Ils forment le stade sexué du cycle vital : des petites méduses. Elles vont développer des organes sexuels et produire des ovules ou des spermatozoïdes.

2.4. L'hydre d'eau douce, Hydra viridis (Hydrozoaire)
Nous allons voir maintenant l'hydre d'eau douce. Elle ne se présente que sous la forme polype et n'engendre donc pas de forme méduse. Celle-ci, munie de longs tentacules, porte un petit bourgeon.
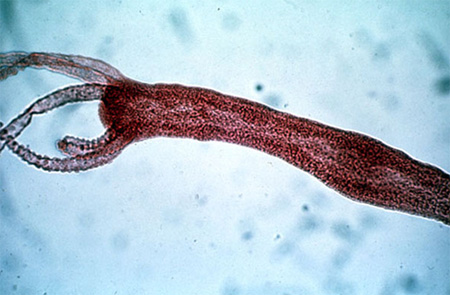
2.4.1. Examen externe
Vous voyez un montage d'hydre colorée au carmin. Retrouvez la forme d'urne, la couronne de tentacules souples et contractiles. A l'intérieur de cette couronne se situe un petit dôme au sommet duquel se trouve la bouche. Le pied de l'hydre lui permet de se fixer sur le fond des ruisseaux ou sur les plantes aquatiques.
2.4.2. Examen interne - 2.4.2.1. Tégument
La composition cellulaire de l'hydre est semblable à celle d’Obelia : elle a un ectoderme périphérique et un endoderme interne, séparés par mince lame de mésoglée.
2.4.2.2. Système locomoteur
Il n’y a pas de locomotion chez l’hydre.
La digestion, la circulation, la respiration et l'excrétion
Se font de la même manière que chez Obelia.
Exercices
1. Voici une coupe microscopique transversale de la paroi de l'Hydre. Repérez-y tous les éléments que vous reconnaissez.
2. Voulez-vous noter le type de symétrie que nous avons rencontré chez les polypes des différentes espèces de Cnidaires que nous avons vu jusqu'ici ?
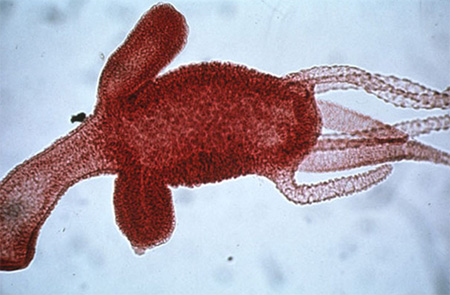
2.4.2.3. Système reproducteur
La reproduction asexuée, par bourgeonnement, est le mode de reproduction usuel pendant les mois chauds de l'année. Les bourgeons proviennent d'une hernie de l'ectoderme et de l'endoderme, hernie qui s'est allongée, qui va se perforer pour former la bouche et qui va produire de petits tentacules. La cavité gastro-vasculaire du bourgeon est en communication avec celle de l'hydre-mère. La jeune hydre se détachera de sa mère par un pincement à la base. Les bourgeons peuvent aussi rester temporairement attachés à la mère pour former une colonie, qui peut compter jusqu'à 15 individus.
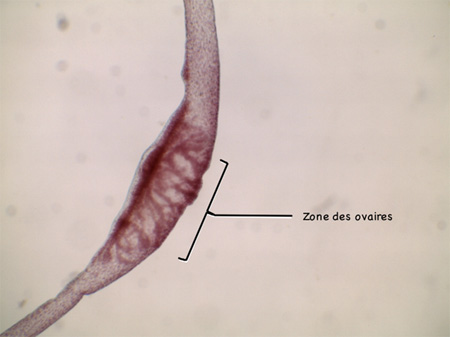
Reproduction sexuée
Mais l'Hydre se reproduit aussi de manière sexuée, principalement en automne ; l'espèce survit à l'hiver en produisant des œufs. L'Hydre est hermaphrodite.
En haut du corps, près des tentacules, se trouvent de petites protubérances. Ce sont les gonades mâles. Plus bas dans le corps se trouve la gonade femelle (les ovaires).
Reproduction sexuée
La figure du syllabus schématise l'ensemble du processus de la reproduction sexuée. Les cellules à l'origine de la formation des gonades mâles et femelles sont les cellules interstitielles qui prolifèrent et forment des hernies sous l'ectoderme. Les cellules sont totipotentes, elles peuvent donc se transformer en n'importe quel type de cellules selon les besoins de l'animal.
Coupe transversale d'Hydre
Voici une coupe transversale d'Hydre au niveau des gonades mâles. Trois gonades mâles sont visibles sous une mince couche de cellules ectodermiques. Arrivés à maturité, les spermatozoïdes percent l'ectoderme et s'échappent dans l'eau.
Coupe transversale
La photo montre aussi une coupe transversale, mais cette fois au niveau de la gonade femelle. On voit nettement qu'une seule cellule interstitielle se développe en ovule. Les autres, dégénérées, servent de substance de réserve. Lorsque l'ovule est mûr, l'ectoderme qui le recouvre éclate mais l'ovule reste ancré dans la paroi.
Reproduction sexuée
L'ovule exposé à l'eau peut être fécondé par un spermatozoïde. La fécondation est croisée bien que l'Hydre soit hermaphrodite. En effet, les spermatozoïdes et les ovules ne sont pas mûrs en même temps sur la même Hydre.
Obelia
La fécondation donne un zygote qui se segmente sur place. Le zygote devient une blastula, puis une gastrula à deux assises cellulaires qui donneront l'ectoderme et l'endoderme. L'assise externe sécrète une gangue protectrice. La gastrula se détache de l'individu mère et tombe au fond de l'eau. Elle peut se transformer immédiatement en jeune polype ou attendre de meilleures conditions du milieu ambiant avant de poursuivre son développement. Chez l'Obélia, la reproduction asexuée est l'oeuvre du polype tandis que la reproduction sexuée est l'oeuvre de la méduse. Chez l'Hydre, les deux formes de reproduction se font par le polype.
Les tentacules de l’hydre verte entourent la bouche. Ils portent des cnidoblastes, qui vont se décharger pour immobiliser les proies. La Daphnie est une telle proie. Les cnidoblastes percent le squelette externe de la Daphnie et injectent un venin qui la paralyse. Les tentacules amènent la Daphnie vers la bouche, qui en se distendant, engloutit le crustacé. La digestion commence. Les cellules endodermiques déversent les enzymes qui digèrent la proie, et absorbent les nutriments. Enfin, les résidus indigestes sont régurgités par la bouche.
Les Hydres
Les Hydres produisent des organes sexuels mâles et femelles. Les testicules se forment à la partie supérieure de la colonne. On voit les spermatozoïdes, très actifs, dans ce testicule. Plus bas sur le corps de l’Hydre, une autre excroissance, plus grosse, représente un ovaire en formation. Il produit un seul gros ovule. Après fécondation, habituellement par un spermatozoïde provenant d’une autre Hydre, l’oeuf s’entourera d’une paroi épaisse, capable de résister aux conditions hivernales.

3. Origine, diversité et évolution
On a décrit environ 9,000 espèces de Cnidaires. Nous n’allons pas voir en détail les données morphologiques et moléculaires qui permettent de retracer l’origine évolutive, la diversité, et les relations de parenté que le groupe des Cnidaires entretient avec les autres embranchements de Métazoaires. Ces questions seront abordées au cours de vos travaux de groupe, ou dans la suite de votre cursus universitaire.
Sachez cependant que la réponse aux trois questions qui nous préoccupent a changé, pour les Cnidaires comme pour la majorité des groupes taxonomiques vus dans ce cours, de manière importante au cours des dix dernières années. Ceci grâce au développement des techniques de séquençage d’ADN et d’expression génique. Il est donc essentiel de consulter des sources d’informations fiables et récentes (telles que les sites de recensement des publications scientifiques en biologie Pubmed ou web of science) si vous désirez, ou devez, approfondir la question. Certains sites sont libres d’accès, d’autres nécessitent une licence. Ces derniers sont accessibles via le site de votre bibliothèque universitaire. Ces sites sont un des principaux moteurs de recherche au niveau mondial pour l’accès en ligne de l’information scientifique vérifiée (au contraire de sites tels que « wikipedia »).
Questions
On peut répondre aux trois questions de la façon suivante :
1) Quelle est l’origine évolutive du groupe taxonomique ?
On a les premières traces de méduses (siphonophores) dans la faune d’Ediacara, il y a 680 millions d’années, mais pas de traces d’organismes à symétrie bilatérale. Une étape majeure de la complexification des Métazoaires concerne pourtant, après l’apparition de la multicellularité, la mise en place de la symétrie bilatérale qui caractérise la grande majorité des Métazoaires. Il est remarquable en effet que la diversité des plans de structure du corps ainsi que la diversité en espèces ait explosé à partir du moment où les animaux ont acquis la bilatéralité et un 3ième feuillet embryonnaire: parmi les Triploblastiques, on retrouve plus de 30 phyla distincts correspondant chacun à un plan de corps différent, et plus de 41 millions d’espèces décrites alors que les Métazoaires plus primitifs (Eponges, Placozoaires, Cténophores et Cnidaires) ne regroupent qu’à peu près 30,000 espèces.
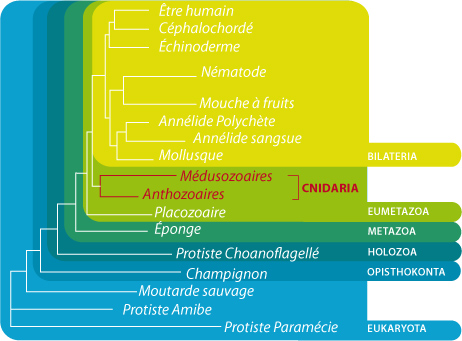
Phylogénie des cnidaires
Différentes phylogénies moléculaires ont maintenant montré que les Anthozoaires sont ancestraux par rapport aux autres groupes de Cnidaires et leurs caractères sont donc ancestraux (on parle de plésiomorphies) pour le groupe, et potentiellement pour le reste des Métazoaires. Les Anthozoaires présentent différentes traces de symétrie bilatérale que vous avez vues dans ce cours en ligne, et que vous verrez plus en détail au TP.
Exercice : lister les caractères à symétrie bilatérale observés chez les Anthozoaires.
Il y aurait donc peut-être un ancêtre commun à la fois aux Cnidaires et aux Métazoaires plus évolués, que certains auteurs appellent « Urbilateria », qui présentait déjà une symétrie bilatérale. Certains groupes de Cnidaires tels que les Hydrozoaires seraient donc revenus secondairement à une morphologie radiaire, sans doute parce que cette dernière est plus adaptée à un mode de vie sessile : pour un organisme fixe, la nourriture et les prédateurs peuvent venir de toutes les directions. En conséquence, certains auteurs ont récemment inclut les Cnidaires parmi les Bilateralia.
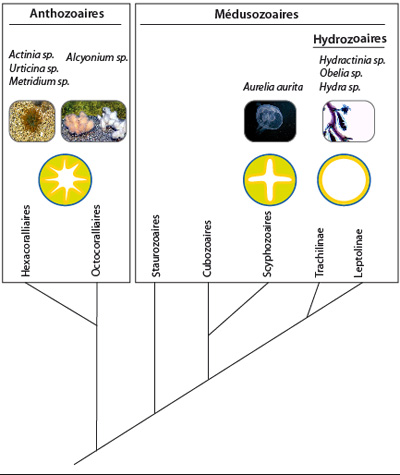
2) Quelle est la diversité existante au sein du groupe taxonomique ?
Deux classes se distinguent parmi les Cnidaires, les Anthozoaires et Medusozoaires, que vous verrez en travaux pratiques. Au sein des Médusozoaires, on retrouve notamment les Hydrozoaires et les Scyphozoaires dont vous avez vu des représentants. Plusieurs critères permettent de ranger ces représentants dans ces classes. Nous en retiendrons un: la structure de la cavité gastro-vasculaire de la forme polype. Les espèces dont la cavité gastro-vasculaire est un sac cylindrique simple sont classées parmi les Hydrozoaires. Celles dont la cavité gastro-vasculaire est quadrilobée parce que l'endoderme fait 4 replis, sont classées parmi les Scyphozoaires. Les espèces dont la cavité gastro-vasculaire est découpée par de nombreux replis de l'endoderme, au moins six paires de replis, sont classées parmi les Anthozoaires.
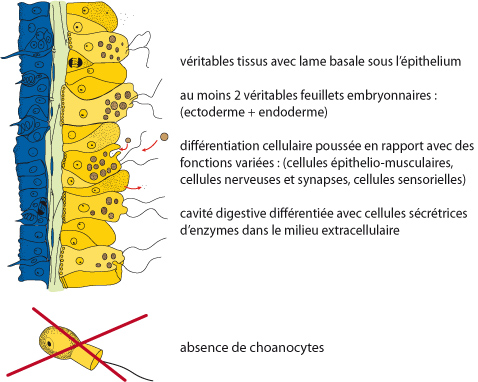
Synapomorphies des cnidaires
Les caractéristiques qui distinguent cet embranchement des Eponges, et qui caractérisent les groupes plus évolués que les Eponges de façon générale, sont: de véritables tissus avec lame basale sous l’epithélium, au moins deux véritables feuillets embryonnaires (ectoderme et endoderme), différenciation cellulaire poussée en rapport avec des fonctions variées (cellules épithélio-musculaires, cellules nerveuses et synapses, cellules sensorielles, ...), cavité digestive différenciée avec cellules sécrétrices d’enzymes digestives dans le milieu extracellulaire, absence des choanocytes. Ces caractères sont communs à tous les Eumétazoaires (ou Bilateralia) ; on parle de synapomorphies pour les Eumétazoaires.
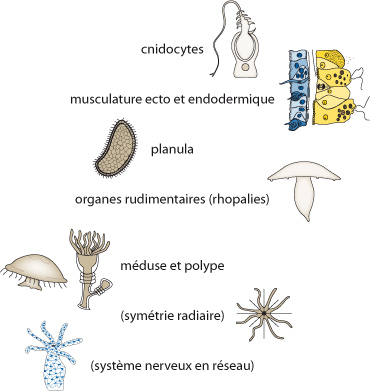
Automorphies des cnidaires
Par ailleurs, une série de caractères distinguent les Cnidaires de tout autre groupe animal ; ce sont les autopomorphies du groupe. Parmi celles-ci, les plus importantes sont les cnidocytes, la musculature d’origine ecto- et endodermiques (et non mésodermique), le stade larvaire de type planula à épiderme cilié avec ectoderme et endoderme et qui est issue de la reproduction sexuée, et l’existence de deux formes, méduse et polype, adaptées à leur mode de vie. Les Cnidaires se ressemblent aussi par l’absence d’appareil circulatoire ou excréteur, un système nerveux très simple disposé en réseau multidirectionnel (pas de synapses chimiques), la symétrie radiaire avec tendance à l’acquisition d’une symétrie bilatérale et par la présence d’organes rudimentaires tels que les rhopalies chez le stade méduse.
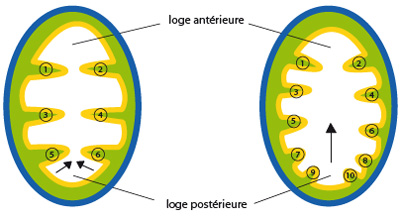
Accroissement métamérique des septa chez un cerianthe
Au sein des Anthozoaires, on peut distinguer les sous-classes des octocoralliaires et des hexacoralliaires qui diffèrent par le pattern de développement des septa mésentériques. Comme nous l’avons vu chez Actinia equina, les Hexacoralliaires, les polypes présentent 6 septa au stade jeune, et ce nombre est variable en cours de vie des polypes et en fonction des espèces. Le nombre de septas peut s’accroître de façon cyclomérique (par ex. Actinia), ou métamérique. C’est le cas chez Cerianthus : la disposition des septa délimite une loge antérieure, une postérieure et 4 latérales. Les nouveaux septa apparaissent toujours dans la loge postérieure, et refoulent vers l'avant les septa préexistants. Chez les Octocoralliaires par contre, on compte 8 septa chez le jeune comme chez l'adulte.
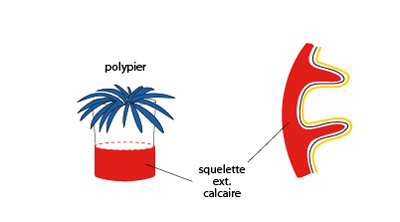
Disposition du squelette des Madrépores
Chez les Madrépores, un hexacoralliaire, le squelette calcaire est une sécrétion de l'ectoderme. Chaque polype sécrète une base et une muraille qui l'engaine à la partie inférieure. Des projections calcaires se développent également en cloisons vers l'intérieur du polype. Ces animaux ont ainsi édifié au cours des ères géologiques les récifs barrières, les îles coralliaires, les atolls. Actuellement, ils s'accroissent de 20 à 40 mm par an. Pour information, notamment pour ceux qui suivront le stage de biologie marine en deuxième quadrimestre, on peut retrouver toute la classification et la description des caractères propres à chaque famille de Cnidaires sur le site suivant : www.ucmp.berkeley.edu/cnidaria/cnidaria.html.
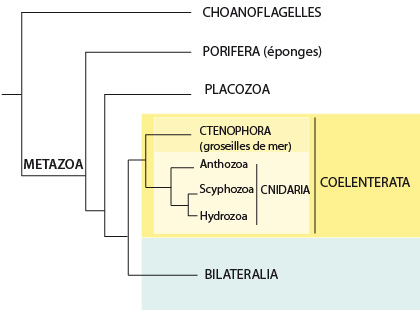
3) Quelles sont les relations de parenté de ce groupe avec les autres groupes étudiés ?
La position des Cnidaires au sein des Métazoaires est encore l’objet de discussions. Il est cependant admis par la majorité des biologistes que les Cnidaires forment un clade avec le groupe des Cténophores ou « groseilles de mer », qu’on appelle « Coelentérés ». Les Coelentérés sont le clade soeur des Bilateralia, qui regroupent tous les animaux que nous verrons dans la suite de ce cours. Les Cnidaires sont donc à la base de la majorité des Métazoaires et cette position primitive explique l’intérêt qu’on porte à l’étude de leurs caractères principaux, tels que cellules musculaires, tissus, organes, épithelia, ...