Le développement des protostomiens
Les Protostomiens constituent un très grand clade d'animaux bilatériens. Il s'agit du groupe frère des deutérostomiens.
Ce groupe a été historiquement caractérisé sur des critères embryologiques, notamment le devenir du blastoderme . Celui-ci donnerait la bouche lors du développement de l'embryon des protostomiens, par opposition aux deutérostomiens où la bouche est néoformée. Il a ensuite été observé que de nombreux protostomiens ont un développement de type deutérostome et les critères embryologiques ont été abandonnés.
Développement de la larve trochophore
La larve trochophore est l’autre caractéristique qui a donné leur nom aux Lophotrochozoaires. Les Annélides et les Mollusques partagent un plan de développement très semblable dans les premiers stades. Nous allons l’illustrer chez Nereis, un Annélide.

Nereis
Nereis que vous voyez ici est un ver Annélide marin. Le Nereis femelle libère ses œufs non fécondés dans l’eau de mer.
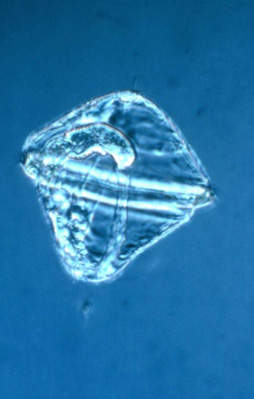
L'oeuf de Nereis
Ces œufs sont sphériques. Ils contiennent bien sûr un noyau, du cytoplasme et une quantité moyenne de vitellus. Comme ils sont moyennement pourvus de vitellus, on les dit 'hétéro-lécithes', par opposition aux œufs télocithes qui en contiennent beaucoup (pensez à l'œuf de la Poule), et aux œufs oligolécithes, qui en possèdent très peu.
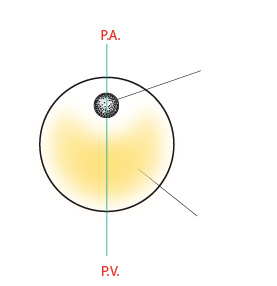
On distingue 2 pôles à cet œuf : le pôle animal (P.A.) et le pôle végétatif (P.V.). Le pôle animal renferme le noyau tandis que le pôle végétatif est plus chargé de vitellus. L'axe passant par les pôles est tracé en vert. Ceci vous rappelle certainement l'oeuf de la Grenouille !
Les oeufs sont fécondés par des spermatozoïdes libérés dans l’eau de mer par un partenaire mâle. L'oeuf fécondé entame alors un processus de segmentation.
La segmentation de l'oeuf
La segmentation est qualifiée de SPIRALE parce que les fuseaux mitotiques ne sont ni horizontaux, ni verticaux, mais obliques par rapport à l'axe de l'œuf qui est déterminé par les pôles animal et végétatif. Lors des deux premières segmentations, c’est peu visible, mais cela devient évident par la suite.
Les embryologistes ont mis au point un codage très pratique (bien qu'un peu compliqué), pour la segmentation spirale que nous allons illustrer ici.
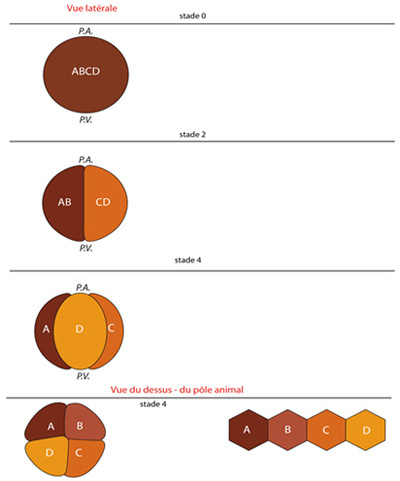
Le zygote que vous voyez latéralement est dénommé ABCD. La première division donne les blastomères AB et CD. La deuxième, les blastomères A, B, C et D. Nous verrons que le blastomère D est appelé à jouer un rôle particulier au cours de l'ontogenèse.
Nous regardons maintenant l'embryon par son pôle animal.
Après la troisième segmentation (stade 8), quatre micromères s'isolent au pôle animal et sont intercalés entre les quatre macromères du pôle végétatif. Les 4 micromères constituent ce que les embryologistes appellent le premier quartette. Les micromères du premier quartette du pôle animal seront appelées 1a, 1b, 1c, 1d. La lettre a minuscule désigne un micromère issu du macromère A majuscule, et rend compte ainsi de son origine. Le coefficient 1 indique le rang: c'est d’ailleurs le PREMIER quartette. Les 4 macromères prennent la désignation 1A, 1B, 1C, 1D (A, B, C, D, en majuscules).
Le stade 16 montre ce qui advient après la quatrième segmentation : les macromères ont donné un deuxième quartette de micromères : 2a, 2b, 2c, 2d. Les macromères sont devenus 2A, 2B, 2C, 2D (les lettres en majuscules). Le premier quartette s'est divisé en même temps, et les cellules sont identifiées par un exposant; on aura 1a1, 1b1, 1c1, 1d1 pour les micromères du premier quartette qui se sont rediviser et 1a2, 1b2, 1c2, 1d2 pour les 4 nouveaux micromères formées au pôle animal. Il y a au total 16 cellules.
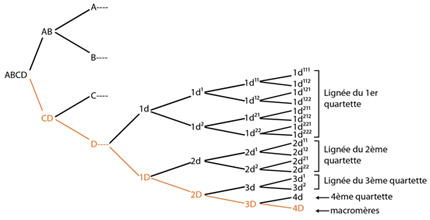
La filiation du blastomère 4D
La segmentation se poursuit : la cinquième, puis la sixième; l'identification est basée sur le même principe, jusqu'à la blastula de 64 blastomères. Nous ne détaillerons pas ces cinquième et sixième segmentations, ce serait très lourd, mais sur cette figure, la filiation du blastomère D y est identifiée jusqu'à la sixième génération. En effet, on peut repérer très tôt les éléments issus d'un blastomère donné, et établir précocement ces filiations.
Nous vous proposons deux questions à titre d'exercice :
Pour quelles raisons pensez-vous que des chercheurs ont passé tant de temps à établir les filiations de la segmentation spirale ?
Comparez la segmentation chez la Grenouille et chez l'Annélide. Citez ce qu'il y a de commun, et ce qui diffère.
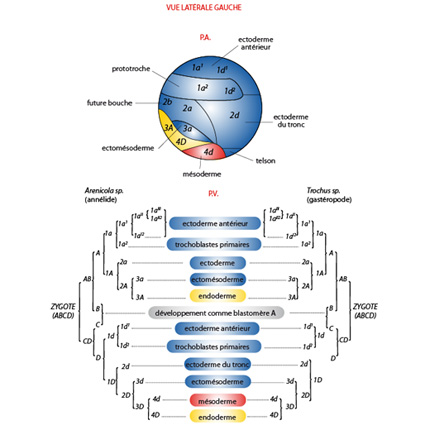
Comparaison des destins des blastomères
Cette figure présente une comparaison des destins des blastomères chez un Annélide, à gauche et un Mollusque, à droite. Elle veut montrer les grandes similitudes qui existent dans les stades embryonnaires précoces entre ces deux Embranchements des Lophotrochozoaires. En particulier, le blastomère 4d du stade 64 cellules sera le seul à former le mésoderme. L’endoderme proviendra de 4 cellules : 3A, 3B, 3C et 4D. Toutes les autres formeront l’ectoderme. Le schéma du dessus montre les territoires présomptifs chez une blastula. Tous les territoires, les feuillets germinatifs, y sont déterminés, ils doivent maintenant se mettre en place.
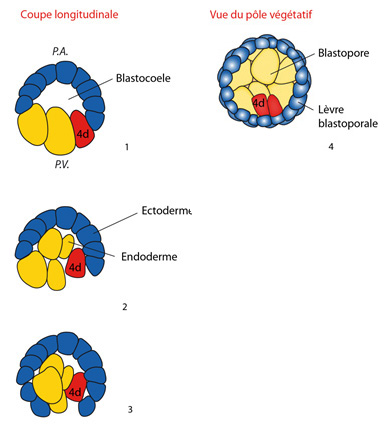
La gastrulation
Chez certains groupes animaux, la gastrulation est fort simple : le pôle végétatif s'enfonce dans le pôle animal, comme un ballon dégonflé dans lequel on donne un coup de poing. Le blastocœle, cavité centrale de la blastula, reçoit cette invagination.
Ici, chez les Annélides, vu la taille des micromères, la blastula est plus souvent massive; la cavité blastocœlienne est fort petite. Regardez l'image 1 qui représente une coupe de blastula, vue latéralement. Alors, sur les images 2 et 3, on voit que la gastrulation se fait par épibolie, c'est-à-dire par étalement des cellules du pôle animal au-dessus de celles du pôle végétatif. L'ensemble des cellules du pôle végétatif se trouve enveloppé par la calotte de micromères. Ces micromères se divisent abondamment et recouvrent les plus grosses cellules passives, qui deviennent internes et représentent l'endoderme, en jaune. Les micromères du revêtement externe constituent l'ectoderme en bleu.
Et le blastomère 4d, qu’en est-il ? Vous le voyez en rouge sur les schémas. D'abord, il est localisé dans la paroi de la blastula, puis il se retrouve coincé entre l'ectoderme et l'endoderme, à l'extrémité postérieure. A ce moment, il se divise et les deux grosses cellules qui en résultent. On le voit sur l'image 4, qui est une vue ventrale de l'embryon. Nous y observons le blastopore, qui est cette espèce de léger enfoncement au fond duquel nous voyons les cellules endodermiques, limité par les lèvres blastoporales. Ces lèvres blastoporales sont le front des micromères bleus qui avancent vers vous et font l'épibolie. De part et d'autre du plan de symétrie de l'embryon, vous voyez les deux cellules filles (en rouge) du blastomère 4d, coincées entre l'ectoderme et l'endoderme et qui représentent le troisième feuillet embryonnaire, le mésoderme.
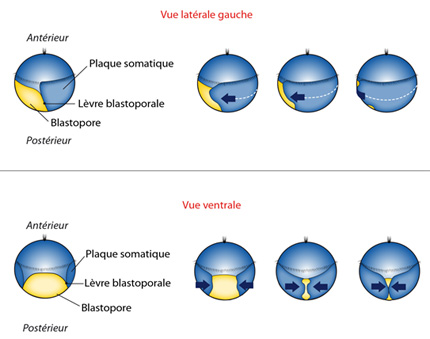
La fermeture du blastophore
A présent, le blastopore va se fermer, regardez l'animation :
- on voit simultanément l'embryon par sa face latérale gauche et par sa face ventrale où se trouve le blastopore.
La calotte ectodermique antérieure, en bleu plus clair, ne participe pas à ce mouvement de fermeture. Remarquez que certaines de ses cellules se sont garnies de cils : une petite touffe au sommet (ce sommet correspond à l'extrémité antérieure), et une couronne à l'équateur. Cet embryon est capable de nager quelque peu avec le battement de ses cils, mais il se laisse surtout emporter par les eaux ; il fait partie du plancton.
En-dessous de l'équateur, la zone ectodermique confinée à la partie dorsale de l'embryon, en bleu foncé, s'appelle « plaque somatique ». Ses bords forment les lèvres du blastopore. Les cellules de la plaque somatique se multiplient et donc la plaque s'étale. Ses lèvres se rapprochent. Vous le voyez durant l'animation :
- les lèvres se rapprochent jusqu'à ce que le blastopore se soit réduit à une simple fente longitudinale, antéro-postérieure. Les lèvres se soudent tout le long de la fente blastoporale, sauf à ses deux extrémités : antérieure et postérieure. Les deux trous qui restent, et au fond desquels on aperçoit encore l'endoderme, donneront naissance à la bouche puis à l'anus.
Ce mode de fermeture du blastopore, par rapprochement et soudure longitudinale ventrale des lèvres blastoporales, porte un nom : c'est la fermeture gastroraphique, du grec 'gaster' : ventre, et 'raphé' : soudure. Ce processus est fondamental pour l'organisation ultérieure du développement.
En guise d'exercice, comparez les modes de fermeture du blastopore chez l'Annélide et chez la grenouille; vous constaterez que c'est très différent comme mouvement. Citez ces différences. Faites bien attention à l'orientation des embryons: les faces antérieures, postérieure, dorsale, ventrale.
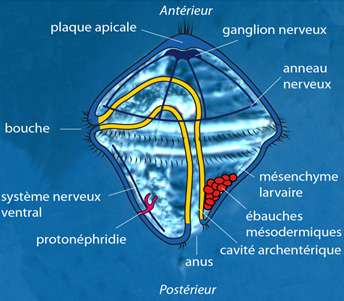
La larve trochophore
Regardez cette figure. Simultanément à la fermeture du blastopore, une série de phénomènes se sont passés. L'endoderme a donné naissance au tube digestif car ses cellules ont ménagé entre elles une cavité allongée : la cavité archentérique.
A la bouche et à l'anus, deux invaginations ectodermiques sont venues à la rencontre des parois antérieure et postérieure endodermiques de la cavité archentérique.
Autre chose : lors du rapprochement des lèvres blastoporales, dans les bourrelets qui avancent, des cordons de cellules se différencient en cellules nerveuses; elles s'enfoncent sous l'ectoderme proprement dit. Les cellules nerveuses constitueront la future chaîne nerveuse longitudinale, qui sera ventrale. Les animaux dotés ainsi d'un système nerveux ventral sont appelés « hyponeuriens » ('hypo' en grec, signifie 'sous'). C'est une caractéristique importante des protostomiens.
En comparaison avec les Annelides hyponeuriens, pouvez-vous justifier le fait que l'on qualifie les Amphibiens, comme les autres Vertébrés, d'épineuriens (en grec epi signifie au-dessus)? Expliquez en comparant les deux modes de développement embryonnaire du système nerveux, chez les hyponeuriens et chez les épineuriens.
L'appareil locomoteur s'est quelque peu diversifié. Une deuxième couronne ciliaire est apparue dans la région équatoriale, en-dessous de la bouche.
La disposition des cils est très variable chez les larves des différentes espèces d'Annélides. Cet appareillage permet de nager, flotter, amener les particules alimentaires vers la bouche... Sous la petite touffe ciliée du sommet, une zone de l'ectoderme s'est épaissie et forme le principal organe sensoriel, la plaque apicale, munie de photorécepteurs, de chémorécepteurs, et d'un organe impliqué dans l'orientation de l’animal.
La coordination de tout ceci est assurée par un ganglion nerveux, situé sous les organes sensoriels apicaux. Une série de nerfs relie ce ganglion à un anneau nerveux équatorial.
En plus, la larve possède quelques structures qui disparaîtront bientôt et que nous ne détaillerons pas, destinées à assurer le maintien général : quelques muscles, des protonéphridies spéciales pour l'excrétion, un mésenchyme de soutien, etc.
Et nous retrouvons la descendance du blastomère 4d ! A présent, les deux cellules filles se sont encore divisées, et constituent deux ébauches mésodermiques pleines, de part et d'autre du tube digestif, au pôle postérieur.
Cette larve trochophore (c'est-à-dire qui porte une roue de cils), on la retrouve plus ou moins modifiée chez un bon nombre d'Invertébrés marins.
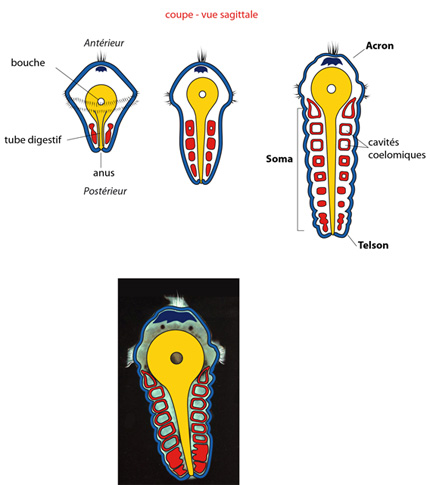
La métamérisation
Maintenant, la métamérisation va apparaître chez la larve trochophore de l’Annélide. Nous la regardons en vue sagittale ; la bouche est dirigée vers vous.
Les ébauches mésodermiques pleines, situées à droite et à gauche de la partie terminale de l'intestin prolifèrent vers l'avant en bandelettes mésoblastiques. La trochophore s'allonge. Puis, alors que la prolifération se poursuit, ces bandelettes se fragmentent à partir de l'avant, en massifs symétriques et réguliers.
Ces massifs se creusent ensuite de cavités : les cavités cœlomiques, limitées par leur paroi. Ce processus de formation des cœlomes à partir de fentes creusées dans le mésoderme s’appelle « schizocœlie » ('schizo' en grec signifie 'fendre'), un processus caractéristique des protostomiens. Finalement, on a un certain nombre de paires de sacs cœlomiques, tous identiques, répartis symétriquement à droite et à gauche du tube digestif, le long de l'axe de la larve. La trochophore métamérisée présente ainsi une structure segmentaire.
Notons deux faits importants.
En arrière des bandelettes mésodermiques métamérisées, il existe une courte zone terminale autour de l'anus, qui ne comporte pas de cavité cœlomique, puisque c'est là que démarrent les proliférations. Cette zone s'appelle le pygidium, aussi appelée telson. De même, dans la région antérieure, existe une zone où les sacs cœlomiques ne se formeront jamais: antérieurement à la bouche et aux couronnes ciliées : c'est le prostomium, ou mieux : acron.
Ainsi, un animal métamérisé comporte, au moins à l'état embryonnaire, une région plus ou moins importante : le soma caractérisé par une succession régulière de sacs cœlomiques pairs, précédé d'une petite région dépourvue de cœlome: l'acron, et suivi d'une autre petite région sans cœlome: le telson.
Sachez que chez certains protostomiens les cavités cœlomiques ne se forment pas par schizocoelie mais c’est le blastocœle (du stade blastula) qui devient la cavité interne du corps. Ceci est le cas par exemple chez les nématodes et les rotifères. Avant on appelait ces groupes d’organismes des « blastocœliens » ou « pseudocoelomates ».
Une question : chez les animaux que nous avons étudiés auparavant, vous vous souvenez sans doute que deux groupes au moins montraient une structure segmentaire répétitive : le strobile des Scyphozoaires, et les Cestodes adultes. Nous n'avons pas parlé alors de métamérisation ou de structures métamériques. Pouvez-vous expliquer pourquoi ?
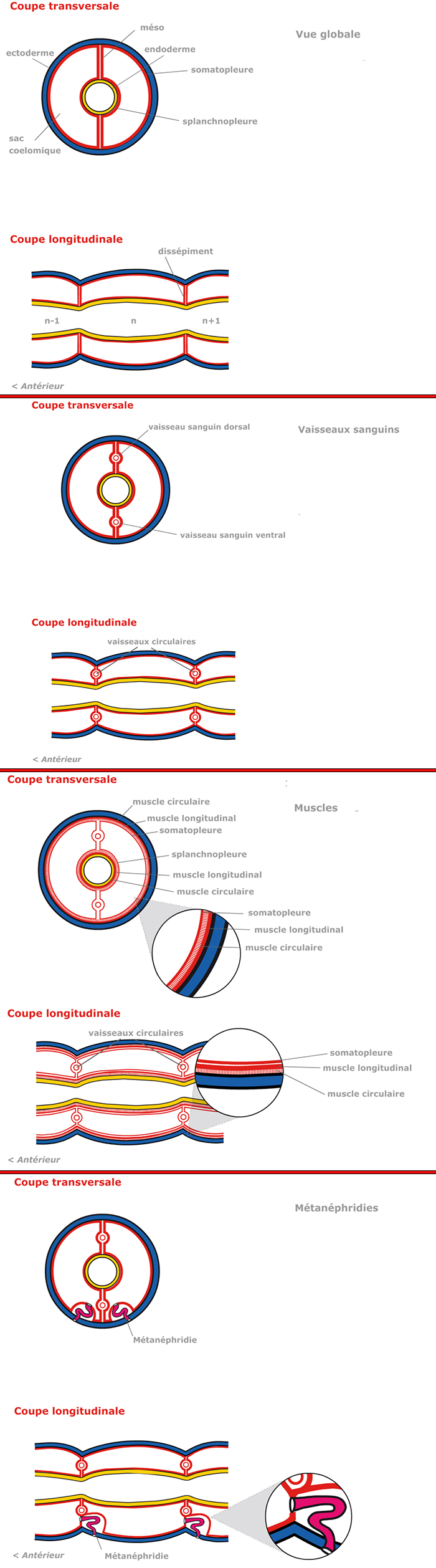
Développement et spécialisation de la métamérisation
Dans la suite du développement de cette larve, l'évolution des sacs cœlomiques va entraîner des conséquences très importantes pour l'anatomie de l'animal.
Tout d'abord, voyez la coupe transversale circulaire. Les sacs cœlomiques accroissent leur volume et occupent tout l'espace compris entre l'ectoderme et l'endoderme. Ainsi, la paroi externe, que l'on appelle somatopleure s'applique étroitement contre l'ectoderme, qui se moule sur elle, d'où l'aspect annelé que prennent les téguments, avec des sillons entre les segments, comme on le voit sur la coupe longitudinale. Ces sillons correspondent exactement aux plans séparant les paires successives de sacs cœlomiques.
« Vue globale »
L'accroissement de volume des sacs cœlomiques a également des conséquences sur l'anatomie interne : les parois médianes des sacs s'accolent dans le plan médian ; elles forment une lame verticale double, le méso (du grec mesos, le milieu), qui suspend l'endoderme intestinal (ce méso est appelé mésentère car situé au milieu de l’intestin, ou enteron en grec). Et l'endoderme intestinal lui-même est emballé dans la paroi cœlomique qui porte le nom de splanchnopleure à ce niveau. Tout naturellement, on distingue un méso dorsal et un méso ventral.
L'accolement des parois cœlomiques se fait aussi dans le sens de la longueur : voyez le schéma de la coupe longitudinale : la paroi antérieure du segment « n+1 » s'applique contre la paroi postérieure du segment « n ». Le tube digestif est alors suspendu transversalement au niveau de chaque intersegment par un disque constitué de 2 couches de mésodermes: c'est le dissépiment.
« Vaisseaux sanguins »
Nous compliquons un peu la situation : l'accolement des parois cœlomiques qui aboutit à la formation des mésos et des dissépiments n'est pas complet.
Dans le cas des mésos, sur la coupe transversale, des espaces tubulaires longitudinaux sont préservés : ils donneront naissance aux vaisseaux sanguins dorsal et ventral, qui du fait même de leur formation, seront continus d'un métamère à l'autre.
Dans le cas des dissépiments, sur la coupe longitudinale, un processus analogue aboutit à la formation de vaisseaux annulaires (ou circulaires), qui font communiquer le vaisseau dorsal et le vaisseau ventral, au niveau de chaque intersegment. Les éléments essentiels du système circulatoire des Annélides, de dérivé mésodermique, présenteront donc une disposition métamérique. Tout comme l'apparence segmentée de l'ectoderme qui est une conséquence directe de la segmentation du mésoderme.
« Muscles »
La musculature constitue un volumineux dérivé du mésoderme.
Situés entre l'ectoderme et la somatopleure, des muscles se sont différenciés aux dépens du mésoderme de la somatopleure. On voit principalement une couche circulaire appliquée contre l'ectoderme, et une couche longitudinale, contre la somatopleure.
Pareillement, la splanchnopleure a édifié une musculature autour de l'intestin ; la constitution de cette musculature intestinale est symétrique de la précédente : une couche circulaire interne et une longitudinale externe.
La somatopleure et la splanchnopleure sont toujours présentes, sous forme de minces feuillets tapissant la cavité cœlomique.
Le plan d'organisation de la musculature est métamérique, aussi parle-t-on de myomérie.
« Métanéphridies »
Le rôle de l'appareil excréteur sera notamment d'établir une communication entre la cavité générale et l'extérieur, de façon à permettre l'évacuation des déchets métaboliques et de l'excès d'eau. Puisque la cavité générale de la grande majorité des Annélides est typiquement cloisonnée transversalement par les dissépiments et longitudinalement par les mésos, c'est chaque sac cœlomique qui devra entrer en communication avec l'extérieur.
Alors, le métamère typique comportera une paire de métanéphridies. La métanéphridie s'ouvre dans la cavité cœlomique par un entonnoir cilié, et un tube conduit à l'extérieur.
Alors, le métamère typique comportera une paire de métanéphridies. La métanéphridie du métamère « n+1 » s'ouvre dans la cavité cœlomique du métamère « n » par un entonnoir cilié, et un tube conduit à l'extérieur au niveau du métamère « n+1 ».
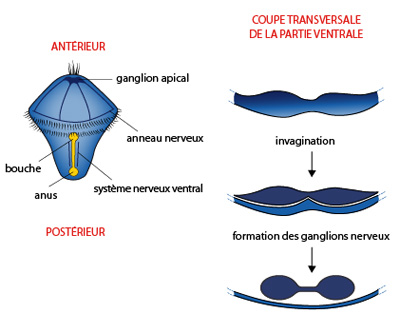
Formation du système nerveux
Rappelons-nous, à la fin de la gastrulation, le blastopore de la larve trochophore était très allongé ; les lèvres blastoporales sont épaissies, et deux bourrelets longitudinaux s'enfoncent sous le tégument pour former deux cordons : la principale ébauche du système nerveux.
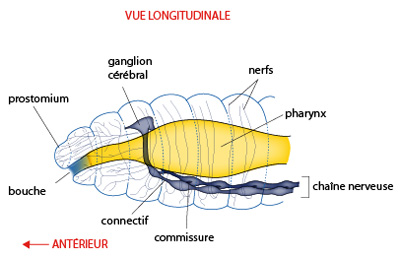
Le système nerveux
Ensuite, probablement sous l'effet inducteur de la métamérisation mésodermique, les cellules ectodermiques composant les cordons nerveux primitifs se rassemblent au milieu de chaque métamère; elles forment ainsi des ganglions nerveux disposés métamériquement, une paire par métamère. Puis, des neurones de chaque ganglion envoient des fibres d'association vers le ganglion voisin, symétrique. Ainsi prennent naissance les commissures transversales, évidemment métamériques.
Donc, le système nerveux prend l'allure d'une échelle dont les barreaux sont les commissures, et dont les montants sont dénommés connectifs. Les ganglions sont bien sûr aux points de jonction entre connectifs et commissures.
Vers l'avant, les connectifs se prolongent jusqu'à rejoindre le système nerveux situé au sommet puisque la chaîne nerveuse ventrale est sous l'intestin, un collier entoure la partie antérieure du tube digestif.
Ce qui se passe à l'autre bout, vers l'arrière dans le telson, n'a que peu d'importance : les rapports de la partie postérieure du corps avec l'environnement n'ont aucune mesure avec ceux de l'extrémité antérieure, sur le plan sensoriel. Il n'y a donc pas de spécialisation particulière du système nerveux dans le telson.

Résumons-nous
A la fin du développement, le métamère typique nous apparaît donc comme un ensemble complexe, constitué d'ectoderme protecteur, du système nerveux dérivé de l'ectoderme, de l'endoderme digestif, ... et d'une variété de descendances mésodermiques: le cœlome et ses parois, les vaisseaux, la musculature, l'appareil excréteur.
Nous vous suggérons de récapituler autrement ce développement : comparez ces métamères de l'Annélide avec les métamères que nous avons vus chez la Grenouille. Cet exercice est assez difficile, mais très riche.